

Расти́тельные кле́тки — эукариотические клетки, однако несколькими своими свойствами они отличаются от клеток остальных эукариот. К их отличительным чертам относят:
- Крупная центральная вакуоль, пространство, заполненное клеточным соком и ограниченное мембраной — тонопластом[1][2]. Вакуоль играет ключевую роль в поддержании клеточного тургора, контролирует перемещение молекул из цитозоля в выделения клетки, хранит полезные вещества и расщепляет отслужившие старые белки и органеллы.
- Есть клеточная стенка, состоящая главным образом из целлюлозы, а также гемицеллюлозы, пектина и во многих случаях лигнина. Она образуется протопластом поверх клеточной мембраны. Она отличается от клеточной стенки грибов, состоящей из хитина, и бактерий, построенной из пептидогликана (муреина).
- Специализированные пути связи между клетками — плазмодесмы[3], цитоплазматические мостики: цитоплазма и эндоплазматический ретикулум (ЭПР) соседних клеток сообщаются через поры в клеточных стенках[4].
- Пластиды, из которых наиболее важны хлоропласты. Хлоропласты содержат хлорофилл, зелёный пигмент, поглощающий солнечный свет. В них осуществляется фотосинтез, в ходе которого клетка синтезирует органические вещества из неорганических. Другими пластидами являются лейкопласты: амилопласты, запасающие крахмал, элайопласты, хранящие жиры и др., а также хромопласты, специализирующиеся на синтезе и хранении пигментов. Как и митохондрии, чей геном у растений содержит 37 генов[5], пластиды имеют собственные геномы (пластомы), состоящие из около 100—120 уникальных генов[6]. Как предполагается, пластиды и митохондрии возникли как прокариотические эндосимбионты, поселившиеся в эукариотических клетках[7].
- Деление клеток (митоз) наземных растений и некоторых водорослей, особенно харовых (Charophyta)[8] и порядка Trentepohliales характеризуется наличием дополнительной стадии — препрофазы. Помимо этого цитокинез у них осуществляется при помощи фрагмопласта — «формы» для строящейся клеточной пластинки[9].
- Мужские половые клетки мхов и папоротниковидных имеют жгутик, схожий со жгутиком сперматозоидов животных[10][11], но у семенных растений — голосеменных и цветковых — они лишены жгутика[12] и называются спермиями.
- Из присущих животной клетке органелл у растительной отсутствуют только центриоли[13].
Сравнение клеток растений и животных
| Признак | Клетки животных | Клетки растений | Исключения |
|---|---|---|---|
| Клеточная стенка | Нет | Есть (целлюлоза) | Нет |
| Тип питания | Гетеротрофные | Автотрофные | Растения-паразиты миксотрофны |
| Пластиды | Нет | Есть | Подземные побеги, растения-паразиты, а также клетки всех тканей, кроме основной лишены хлорофилла |
| Центриоли | Есть | Нет | Нет |
| Центральная вакуоль | Нет | Есть | У растений отсутствует в мёртвых и др. специализированных клетках |
| Основное запасное питательное вещество |
Гликоген | Крахмал | Нет |
| Целостные реакции клетки (пиноцитоз, эндоцитоз, экзоцитоз, фагоцитоз) |
Есть | Нет | Нет |
Основные типы растительных клеток
Паренхимные клетки

Паренхимные клетки — это клетки, размеры которых во всех направлениях одинаковы или длина немного больше ширины[14]. Паренхиму растений называют также основной тканью[15].
Клетки паренхимы образуют однородные скопления в теле растения, заполняют пространства между другими тканями, входят в состав проводящих и механических тканей. Они могут выполнять различные функции: ассимиляционную, выделительную и др. Приспособленность паренхимных клеток к различным функциям обусловлена их функциональной специализацией протопластов. Присутствие в паренхиме (особенно рыхлой) межклетников определяет её участие в газообмене. Живые паренхимные клетки способны к делению; в паренхиме закладывается феллоген, а у растений с атипичным приростом в толщину — камбий (корнеплоды свёклы, некоторые лианы)[15].
Прозенхимные клетки
Прозенхимные клетки — это вытянутые (длина во много раз превышает ширину) и заострённые на концах (в отличие от паренхимы) клетки, различные по происхождению и функциям. Между прозенхимой (тканью, образованной прозенхимными клетками) и паренхимой имеются переходы, например, колленхима и лопастные ветвистые клетки мезофилла в листьях канны и др. растений[16].
Деление растительных клеток

- N — ядро,
- V — вакуоль,
- PPB — препрофазная лента,
- MTN — начало скопления микротрубочек,
- NEB — распад ядерной оболочки в начале прометафазы.
У растительных клеток имеется уникальная дополнительная фаза митоза — препрофаза. Она предшествует профазе и включает два основных события:
- Образование препрофазной ленты — кольца из микротрубочек, расположенного под плазматической мембраной[18];
- Начало скопления микротрубочек около ядерной оболочки[19].
В остальном митоз растительных клеток проходит так же, как у остальных эукариот, только цитокинез у них протекает с использованием специальных структур — фрагмопласта (у высших растений и некоторых водорослей), фикопласта (у нек. других водорослей) и др.
Клеточная стенка
Клеточная стенка имеется не только у растительных клеток: она есть у грибов и бактерий, но только у растений она состоит из целлюлозы (исключением являются грибоподобные организмы оомицеты, чья клеточная стенка также состоит из целлюлозы[20]).
Структура и химический состав
Клеточная стенка образуется из клеточной пластинки, причём сначала формируется первичная, а затем вторичная клеточная стенка. Строение клеточной стенки двух этих типов напоминает устройство железобетонных блоков, в которых присутствует металлический каркас и связующее вещество — цемент. В клеточной стенке каркасом являются пучки молекул целлюлозы, а связующим веществом служат гемицеллюлоза и пектины, которые образуют матрикс клеточной стенки. Эти вещества транспортируются во время роста клеточной пластинки из комплекса Гольджи к плазматической мембране, где пузырьки сливаются с ней и посредством экзоцитоза выбрасывают содержимое наружу[21].
Помимо указанных веществ, в оболочке одревесневших клеток содержится лигнин, повышающий их механическую прочность и понижающий водонепроницаемость. Кроме того, в оболочке клеток некоторых специализированных тканей могут накапливаться гидрофобные вещества: растительные воска, кутин и суберин, откладывающийся на внутренней поверхности стенок клеток пробки и составляющий пояски Каспари[22].
Первичная и вторичная клеточные стенки
Первичная клеточная стенка содержит до 90% воды и характерна для меристематических и малодифференцированных клеток. Эти клетки способны изменять свой объём, но не за счёт растяжения целлюлозных фибрилл, а смещения относительно друг друга этих фибрилл[23].
Некоторые клетки, например, мезофилла листа, сохраняют первичную оболочку и по достижении нужных размеров перестают откладывать в неё новые вещества. Однако у большинства клеток этот процесс не прекращается, и между плазматической мембраной и первичной оболочкой у них откладывается вторичная клеточная стенка. Она имеет принципиально схожее с первичной строение, но содержит значительно больше целлюлозы и меньше воды. Во вторичной стенке обычно различают три слоя — наружный, самый мощный средний и внутренний[23].
Поры
Во вторичной стенке имеется большое количество пор[23]. Каждая пора представляет собой канал в том месте клеточной оболочки, в котором над первичным поровым полем не откладывается вторичная оболочка[24]. Первичное поровое поле — это небольшой участок тонких смежных стенок двух клеток, состоящий из первичной оболочки и клеточной пластинки, пронизанный плазмодесмами[25]. Поры возникают парно в смежных клетках соседних клеток и разделены замыкающей трёхслойной[25] плёнкой (поровой мембраной)[24]. Различают поры:
- Простые поры представляют собой каналы во вторичной оболочке паренхимных клеток и склереид, имеющие одинаковую ширину на всем протяжении.
- Окаймлённые поры — это поры, окаймление которых составляет куполообразно приподнятая над поровой мембраной вторичная оболочка. В плане такая пора имеет вид двух окружностей, наружная из которых соответствует окаймлению, а внутренняя — отверстию, открывающемуся в полость клетки. Характерны для водопроводящих элементов, представленных мёртвыми клетками.
- Полуокаймлённые поры — пара пор, одна из которых — простая, другая — окаймлённая. Образуется в смежных стенках трахеид хвойных и паренхимных клеток древесинных лучей.
- Слепые поры представляют собой каналы во вторичной оболочке только одной из двух соседних клеток, такие поры не функционируют.
- Ветвистые поры — поры, разветвлённые на одном из концов вследствие слияния двух или нескольких простых пор в процессе утолщения вторичной оболочки.
- Щелевидные поры — поры с отверстиями в виде косой щели; образуются в клетках прозенхимы, например, волокнах древесины[26].
Плазмодесмы

1 — клеточная стенка
2 — плазмалемма
3 — десмотубула
4 — эндоплазматический ретикулум
5 — белки плазмодесмы
Плазмодесма — это тончайший тяж цитоплазмы, канал, связывающий протопласты соседних клеток[27]. Эти каналы по всей длине выстланы плазматической мембраной. Через плазмодесмы проходит полая структура — десмотубула, через неё элементы ЭПР соседних клеток сообщаются между собой.
Через плазмодесмы осуществляется свободный транспорт веществ. Предполагают, что ситовидные поля флоэмы также представляют собой крупные плазмодесмы[28].
Внутреннее пространство растения, объединяющее все протопласты, связанные посредством плазмодесм, называют симпластом, соответственно, транспорт через плазмодесмы называют симпастическим[29].
Функции
Клеточные стенки растений выполняют следующие функции:
- обеспечение возможности тургора (не будь её, внутриклеточное давление разорвало бы клетку);
- роль наружного скелета (то есть придаёт форму клетке, определяет рамки её роста, обеспечивает механическую и структурную поддержку);
- запасает питательные вещества[29];
- защита от внешних патогенов.
Органеллы
Пластиды

Пластиды — органеллы растительной клетки, состоящие из белковой стромы, окружённой двумя липопротеидными мембранами. Внутренняя из них образует внутрь выросты (тилакоиды, или ламеллы)[30].
Пластиды, как и митохондрии, являются самовоспроизводящимися органеллами и имеют собственный геном — пластом, а также рибосомы.
У высших растений все пластиды происходят от общего предшественника — пропластид, которые развиваются из двумембранных инициальных частиц.
Пластиды присущи исключительно растениям[31]. Различают три основных типа пластид:
- Лейкопласты. Эти пластиды не содержат никаких пигментов, внутренняя мембранная система, хотя и присутствует, но развита слабо. Разделяют амилопласты, запасающие крахмал, протеинопласты, содержащие белки, элайопласты (или олеопласты), запасающие жиры. Этиопласты — это бесцветные пластиды растений, которые выращивали без освещения. При наличии света они легко превращаются в хлоропласты[32].
- Хромопласты — пластиды жёлто-оранжевого цвета, обусловленного наличием в них пигментов каротиноидов: каротина, ксантофилла, лютеина, зеаксантина и др. Образуются из хлоропластов при разрушении в них хлорофилла и внутренних мембран[33]. Кроме того, хромопласты мельче хлоропластов по размерам. Каротиноиды присутствуют в хромопластах в виде кристаллов или растворёнными в каплях жира (такие капли называют пластоглобулами). Биологическая роль хромопластов до сих пор неясна[34].
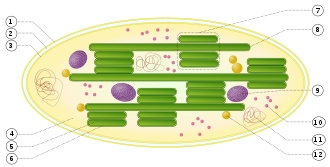
1. наружная мембрана
2. межмембранное пространство
3. внутренняя мембрана (1+2+3: оболочка)
4. строма (жидкость)
5. тилакоид с просветом (люменом) внутри
6. мембрана тилакоида
7. грана (стопка тилакоидов)
8. тилакоид (ламелла)
9. зерно крахмала
10. рибосома
11. пластидная ДНК
12. пластоглобула (капля жира)
- Хлоропласты — пластиды в виде двояковыпуклой линзы, окружённые оболочкой из двух липопротеидных мембран. Внутренняя из них образует длинные выросты в белковую строму — тилакоиды стромы и более мелкие, расположенные стопками тилакоиды гран, соединённые между собой тилакоидами стромы. С белковым слоем мембран тилакоидов связаны пигменты: хлорофилл и каротиноиды. В хлоропластах осуществляется фотосинтез. Первичный крахмал, синтезированный хлоропластами, откладывается в строме между тилакоидами[35].
Гигантские хлоропласты водорослей, присутствующие в клетке в единственном числе, называются хроматофорами. Их форма может быть очень разнообразной[31].
Вакуоли
Вакуоль — полость в клетке, заполненная клеточным соком и окружённая мембраной — тонопластом. Вещества, содержащиеся в клеточном соке, определяют величину осмотического давления и тургор клеточной оболочки.
Вакуоли образуются из провакуолей — небольших мембранных пузырьков, отшнуровывающихся от ЭПР и комплекса Гольджи. Потом пузырьки сливаются, образуя более крупные вакуоли. Только у старых клеток все вакуоли могут сливаться в одну гигантскую центральную вакуоль, обычно же клетка, помимо центральной вакуоли, содержит мелкие вакуоли, наполненные запасными веществами и продуктами обмена[36].
Вакуоли выполняют в клетке следующие основные функции:
- создание тургора;
- запасание необходимых веществ;
- отложение веществ, вредных для клетки;
- ферментативное расщепление органических соединений (это сближает вакуоли с лизосомами)[37].
Включения растительных клеток
- трофические включения:
- крахмальные зёрна;
- белковые гранулы (в гиалоплазме, пластидах, ЭПР, вакуолях, ядре). Чаще всего белковые отложения обнаруживают в виде алейроновых зёрен, представляющих собой наполненные белком обезвоженные вакуоли.
- липидные капли — мощный источник энергии. Энергетическая ценность липидов вдвое выше, чем у белков или углеводов, поэтому содержащие их ткани или семена могут иметь меньшую массу и размеры.
- включения, не имеющие энергетической ценности, как правило, отходы жизнедеятельности. Чаще всего встречаются кристаллы оксалата кальция[38].
Примечания
- ↑ JA Raven (1997) The vacuole: a cost-benefit analysis. Advances in Botanical Research 25, 59–86
- ↑ RA Leigh and D Sanders (1997) Advances in Botanical Research, Vol 25: The Plant Vacuole. Academic Press, California and London. ISBN 0-12-441870-8
- ↑ Oparka, KJ (1993) Signalling via plasmodesmata-the neglected pathway. Seminars in Cell Biology 4, 131–138
- ↑ Hepler, PK (1982) Endoplasmic reticulum in the formation of the cell plate and plasmodesmata. Protoplasma 111, 121–133
- ↑ Anderson S, Bankier AT, et al. (1981) Sequence and organization of the human mitochondrial genome. Nature 290, 4–65
- ↑ L Cui, N Veeraraghavan, et al. (2006) ChloroplastDB: the chloroplast genome database. Nucleic Acids Research, 34, D692-696
- ↑ L. Margulis (1970) Origin of eukaryotic cells. Yale University Press, New Haven
- ↑ Lewis, LA, McCourt, RM (2004) Green algae and the origin of land plants. American Journal of Botany 91, 1535–1556
- ↑ López-Bautista, JM, Waters, DA and Chapman, RL (2003) Phragmoplastin, green algae and the evolution of cytokinesis. International Journal of Systematic and Evolutionary Microbiology 53, 1715–1718
- ↑ Manton, I. and Clarke, B. (1952) An electron microscope study of the spermatozoid of Sphagnum. Journal of Experimental Botany 3, 265–275
- ↑ D.J. Paolillo, Jr. (1967) On the structure of the axoneme in flagella of Polytrichum juniperinum. Transactions of the American Microscopical Society, 86, 428–433
- ↑ PH Raven , Evert RF, Eichhorm SE (1999) Biology of Plants, 6th edition. WH Freeman, New York
- ↑ Билич, Крыжановский, 2009, с. 136.
- ↑ Лотова, Нилова, Рудько, 2007, с. 58.
- ↑ 1 2 Паренхима // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- ↑ Прозенхима — статья из Биологического энциклопедического словаря
- ↑ Dhonukshe, P., Mathur, J., Hülskamp, M., Gadella, TWJ. Microtubule plus-ends reveal essential links between intracellular polarization and localized modulation of endocytosis during division-plane establishment in plant cells (англ.) // BMC Biology : journal. — 2005. — Vol. 3. — P. 11. — doi:10.1186/1741-7007-3-11. — PMID 15831100. — PMC 1087477.
- ↑ Ambrose J. C., Cyr R. J. Mitotic spindle organization by the preprophase band (англ.) // Molecular Plant : journal. — 2008. — Vol. 1, no. 6. — P. 950—960. — doi:10.1093/mp/ssn054. — PMID 19825595. Архивировано 15 апреля 2013 года.
- ↑ Hoshino H., Yoneda A., Kumagai F., Hasezawa S. Roles of actin-depleted zone and preprophase band in determining the division site of higher-plant cells, a tobacco BY-2 cell line expressing GFP-tubulin (англ.) // Protoplasma : journal. — 2003. — Vol. 222, no. 3—4. — P. 157—165. — doi:10.1007/s00709-003-0012-8. — PMID 14714204.
- ↑ Гл. ред. М. С. Гиляров; Редкол.: А. А. Бабаев, Г. Г. Винберг, Г. А. Заварзин и др. Оомицеты // Биологический энциклопедический словарь.» — 2-е изд., исправл. — М.: Сов. Энциклопедия. — 1986.
- ↑ Билич, Крыжановский, 2009, с. 140.
- ↑ Билич, Крыжановский, 2009, с. 146—147.
- ↑ 1 2 3 Билич, Крыжановский, 2009, с. 143.
- ↑ 1 2 Лотова, Нилова, Рудько, 2007, с. 65.
- ↑ 1 2 Лотова, Нилова, Рудько, 2007, с. 59.
- ↑ Лотова, Нилова, Рудько, 2007, с. 65—66.
- ↑ Лотова, Нилова, Рудько, 2007, с. 62.
- ↑ Билич, Крыжановский, 2009, с. 144.
- ↑ 1 2 Билич, Крыжановский, 2009, с. 148.
- ↑ Лотова, Нилова, Рудько, 2007, с. 62—63.
- ↑ 1 2 Билич, Крыжановский, 2009, с. 149.
- ↑ Билич, Крыжановский, 2009, с. 154.
- ↑ Лотова, Нилова, Рудько, 2007, с. 90.
- ↑ Билич, Крыжановский, 2009, с. 152.
- ↑ Лотова, Нилова, Рудько, 2007, с. 89.
- ↑ Лотова, Нилова, Рудько, 2007, с. 17—18.
- ↑ Билич, Крыжановский, 2009, с. 157—160.
- ↑ Билич, Крыжановский, 2009, с. 163—167.
Литература
- Билич Г.Л., Крыжановский В.А. Биология. Полный курс: В 4 т. — издание 5-е, дополненное и переработанное. — М.: Издательство Оникс, 2009. — Т. 1. — 864 с. — ISBN 978-5-488-02311-6.
- Лотова Л. И., Нилова М.В., Рудько А.И. Словарь фитоанатомических терминов: учебное пособие. — М.: Издательство ЛКИ, 2007. — 112 с. — ISBN 978-5-382-00179-1.
| В библиографических каталогах |
|---|
| Органеллы |  | ||||
|---|---|---|---|---|---|
| Клеточная стенка |
| ||||
| Деление растительной клетки |
| ||||
Обычно почти сразу, изредка в течении часа.