
| Красный огненный муравей | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 Представители красных огненных муравьёв Solenopsis invicta | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Без ранга: Тип: Подтип: Надкласс: Класс: Подкласс: Инфракласс: Надотряд: Hymenopterida Отряд: Подотряд: Инфраотряд: Надсемейство: Семейство: Подсемейство: Триба: Род: Вид: Красный огненный муравей |
||||||||||
| Международное научное название | ||||||||||
| Solenopsis invicta Buren, 1972 | ||||||||||
| Исторический ареал | ||||||||||
 |
||||||||||
| ||||||||||
Красный огненный муравей[1][2][3][4], Муравей огненный импортный красный[5] (лат. Solenopsis invicta, англ. Red imported fire ant, RIFA) — один из самых опасных в мире инвазивных видов муравьёв, обладающий сильным ядом, который они впрыскивают в кожу с помощью жала из железы находящейся в задней части брюшка. Для людей их ужаления очень болезненны и могут быть смертельно опасными для лиц, страдающих от аллергии. Кроме того, вторжение этих муравьев на новые территории часто приводит к существенному ущербу для местной природы и экономическим потерям. Относится к группе огненных муравьёв[6].
История
Исторический ареал огненных муравьёв находится в центральных районах Южной Америки. В 1930-е годы их колонии в ходе морских перевозок были случайно завезены в США. Непреднамеренная интродукция произошла через морской порт Мобил (Алабама) в штате Алабама[7][8], где торговые суда из Бразилии вставали в доки для выгрузки контейнеров, некоторые из которых оказались заражены муравьями. Начав своё вторжение с Алабамы, огненные муравьи заселили практически все южные штаты США от Техаса до Мэриленда, а также северо-восточные районы Мексики. В своей новой области расселения они начали оказывать существенное негативное влияние на местную природу и стали причиной значительных экономических потерь[9][10][11][12]. С 1990-х годов их отмечают в Калифорнии (на западе США) и в Нью-Мексико (на юго-западе США), куда они попали, возможно, через суда, заходившие в порты Калифорнии[6].
Схожими путями эти муравьи случайно были завезены в Старый Свет, в Австралию в 2001 году[13] и Новую Зеландию[9].
В сентябре 2004 года красные огненные муравьи зафиксированы на острове Тайвань, где ещё с десяток лет назад появился другой огненный муравей — Solenopsis geminata. После сообщений об опасных последствиях ужаления муравьями жителях острова была предпринята массовая кампания по уничтожению нового инвазивного вида. Однако её эффективность оказалась ограниченной, и в итоге муравьи обосновались также и здесь. В Китае интродукция огненных муравьёв официально была признана в январе 2005 года, хотя сообщения в прессе появлялись ещё с середины 2004 года (газеты Гонконга, включая Appledaily, Mingpao, Hong Kong Economic Times, Singtao и Takungpao). Указывались Шэньчжэнь и Учуань в Чжаньцзяне в провинции Гуандун, а также Гонконг[14].
В июле 2005 года колония огненных муравьёв впервые была отмечена в городе Манила и в Кавите (Филиппины). В 2007 году их нашли уже в Бикольском регионе[9].
За последние полвека было создано несколько специализированных лабораторий и центров по исследованию огненных муравьёв, защищено около 100 диссертаций по темам, непосредственно связанным с этим опасным инвазивным видом, опубликовано более 3000 статей только по одному этому виду муравьёв[15].
Распространение

Родиной огненного муравья является тропическая Южная Америка (Бразилия, Аргентина и Перу)[9]. В современный инвазивный ареал входят США, Мексика, острова Карибского бассейна (Американские Виргинские острова, Антигуа и Барбуда, Багамы, Каймановы острова, Пуэрто-Рико, Теркс и Кайкос, Тринидад и Тобаго), Австралия, Малайзия, Сингапур, Тайвань, Филиппины, Китай (Гуандун, Макао). В США огненные муравьи распространились по штатам Алабама, Арканзас, Аризона, Виргиния, Джорджия, Калифорния, Кентукки, Луизиана, Мэриленд, Миссисипи, Оклахома, Северная Каролина, Теннесси, Техас, Нью-Мексико, Флорида, Южная Каролина[16].
Описание



Размеры красных огненных муравьёв варьируются от мелких до средних, что делает их полиморфными. Размеры рабочих составляют от 2,4 до 6,0 мм[17]. Основная окраска тела рыжевато-коричневая (самцы чёрные). У самок и рабочих 10-члениковые усики с 2-члениковой булавой на конце[18]. Голова рабочих имеет размеры от 0,66 до 1,41 мм в длину и от 0,65 до 1,43 мм в ширину. У более крупных рабочих (как и у солдат) их голова имеет размеры от 1,35 до 1,40 мм и от 1,39 до 1,42 мм в ширину. Скапус усиков имеет размеры от 0,96 до 1,02 мм, а длина груди составляет от 1,70 до 1,73 мм[18][19]. Голова шире за глазами, затылочные доли округлые. Скапус у крупных рабочих не выходит за пределы затылка на один-два диаметра скапуса. У средних рабочих скапус достигает затылочного края и превышают заднюю границу у самых мелких рабочих. У мелких и средних рабочих бока головы более эллиптические. Голова мелких рабочих спереди шире, чем сзади[18]. У крупных рабочих переднеспинка не имеет угловатых плеч. Промезонотум выпуклый, основание проподеума округлое и также выпуклое. Горизонтальная и покатая поверхности имеют одинаковую длину. Шов промезонотума у более крупных рабочих либо сильный, либо слабый. Петиоль имеет толстую чешуйку. Постпетиоль большой и широкий, а у более крупных рабочих он шире своей длины. Постпетиоль имеет тенденцию быть менее широким спереди и более широким сзади[18].
Скульптура хитиновой поверхности тела очень похожа на таковую у чёрного огненного муравья Solenopsis richteri[18]. Пунктуры находятся там, где расположены волоски, и они часто имеют удлиненную форму на дорсальной и вентральной частях головы. На груди имеются бороздки, но они менее выгравированы и с меньшим количеством точек, чем у S. richteri. На петиоле пунктуры расположены по бокам. Постпетиоль, если смотреть сверху, имеет сильную шагренированную скульптуру с отчётливыми поперечными точечными штрихами. Бока покрыты глубокими пунктурами, где они кажутся меньшими, но более глубокими. У S. richteri, точки более крупные и неглубокие. Это придает поверхности более матовый вид[18]. Волосистость похожа на таковую S. richteri. Эти волоски отстоящие и различаются по длине, кажутся длинными с каждой стороны переднеспинки и мезонотума; на голове длинные волосы расположены продольными рядами. На чешуйке петиоля многочисленные прижатые волоски опушения; наоборот у S. richteri эти волосы редкие[18]. Рабочие выглядят красными и несколько желтоватыми с коричневым или полностью чёрным брюшком[20]. У более крупных рабочих иногда наблюдаются светлые пятна на брюшке, где они не так ярко окрашены, как у S. richteri. Эти пятна обычно занимает небольшую часть первого тергита брюшка. Грудь одноцветная, от светло-красновато-коричневого до темно-коричневого цвета. Ноги и тазики обычно слегка затемнены. Голова имеет постоянный цветовой рисунок у крупных рабочих, затылок и темя коричневые. Остальные части головы, включая переднюю часть, щёки и центральную часть наличника, желтоватые или желтовато-коричневые. Передние края щёк и мандибулы темно-коричневые. Скапус и жгутик усика варьируются от того же цвета, что и голова, или того же оттенка, что и затылок. Светлые области головы у рабочих малого и среднего размера ограничены только лобной областью, при этом присутствует тёмная отметина, напоминающая стрелу или ракету. Иногда в гнезде могут быть рабочие нескольких разных цветов. Например, рабочие могут быть значительно темнее, а брюшные пятна могут полностью отсутствовать или быть темно-коричневыми[18].
У маток длина головы 1,27—1,29 мм (ширина 1,32—1,33 мм). Длина скапуса усиков маток от 0,95—0,98 мм, а длина их груди 2,60—2,63 мм. Стебелёк, соединяющий грудку с брюшком двухчлениковый (петиоль + постпетиоль). Постпетиоль имеет прямые стороны, которые не вогнуты, в отличие от S. richteri, у которых они вогнуты. Боковые части петиоля пунктированные. Боковые стороны постпетиоля матовые, с пунктурами, но шероховатости не видно. Передняя часть дорзума шагреневая, средняя и задняя области с поперечными точечными штрихами. Все эти области имеют торчащие волоски. Передние части как петиоля, так и постпетиоля имеют прижатое опушение, которое также видно на проподеуме. Окраска матки похожа на рабочих: брюшко темно-коричневое, ноги, скапус и грудь светло-коричневые с темными пестринами на среднеспинке. Голова желтоватая или желтовато-коричневая вокруг центральных областей, затылок и жвалы того же цвета, что и грудь, а жилки крыльев варьируются от бесцветных до бледно-коричневых. Самцы крылатые, внешне похожи на S. richteri, но верхние края чешуйки петиоля более вогнутые. У обоих видов сильно выступают дыхальца петиоля и постпетиоля. Всё тело самца однотонно-чёрное, но усики беловатые. Как и у матки, жилки крыльев бесцветные или бледно-коричневые[18].
Строят земляные муравейники с холмиками высотой и диаметром до 50 см. В гнёздах обнаружены многочисленные мирмекофилы, например, клещи Gaeolaelaps invictianus[21]. Важную роль в жизни муравьёв играет яд, так как он используется для поимки добычи или для защиты[22].
Расплод
Яйца мелкие, овальной формы, сохраняют один и тот же размер около недели. Через одну неделю яйцо принимает форму эмбриона и превращается в личинку, когда яичная оболочка удаляется[23]. Личинки имеют размер около 3 мм[24]. По внешнему виду они похожи на личинок S. geminata, но их можно отличить по покровам с шипиками на вершине дорсальной части задних сомитов. Волоски на теле имеют размеры от 0,063 до 0,113 мм с зубчатым кончиком. Оба усика имеют две или три сенсиллы. Верхняя губа мелкая и с двумя волосками на передней поверхности размером 0,013 мм. Максилла имеет склеротизованную полосу между кардо и стипесом. На губе также есть небольшая склеротизованная полоса[24]. Известно, что лабиальные железы вырабатывают или выделяют белковое вещество с высоким уровнем пищеварительных ферментов, включая протеазы и амилазы, которые функционируют как внекишечное переваривание твердой пищи. Средняя кишка также содержит амилазы, ротеазы и упазы. Узкие клетки в её резервуаре практически не участвуют в секреции[25]. Куколки напоминают взрослых особей любой касты, за исключением того, что их ноги и усики плотно прижаты к телу. Они выглядят белыми, но со временем куколки становятся темнее, когда они почти готовы к созреванию[26].
Четыре личиночных возраста были описаны на основе отличительных морфологических признаков[23][27][28]. Личинки малых и крупных рабочих невозможно различить до последнего возраста, когда становятся очевидными различия в размерах[28]. При окукливании становится более заметной разница в ширине головы между кастами. Личинки маток и самцов крупнее личинок рабочих и имеют дискретные морфологические различия в ротовом аппарате[28]. Личинки четвёртого возраста самцов и маток можно отличить по их относительной форме и окраске тела[28].
Поведение и экология

Красные огненные муравьи чрезвычайно выносливы и приспособились бороться как с наводнениями, так и с засухой. Если муравьи чувствуют повышение уровня воды в своих гнёздах, они соединяются вместе и образуют плавучий шар или плот с рабочими снаружи и королевой внутри[29][30][31]. Расплод транспортируется на самую высокую поверхность[32]. Они также используются в качестве основы плота, за исключением яиц и более мелких личинок. В некоторых случаях рабочие могут намеренно снять всех самцов с плота, в результате чего самцы утонут. Срок службы плота может достигать 12 дней. Муравьи, попавшие в ловушку под водой, спасаются, поднимаясь на поверхность с помощью пузырей, которые собираются из подводного субстрата[32]. Из-за большей уязвимости для хищников красные огненные муравьи значительно более агрессивны при сплаве. Рабочие, как правило, вводят более высокие дозы яда, что снижает угрозу нападения других животных. Из-за этого такие муравьиные плоты потенциально опасны для тех, кто с ними сталкивается[33].
У красного огненного муравья отмечено поведение некрофореза. Рабочие выбрасывают несъеденную пищу и другие подобные отходы подальше от гнезда. Активный компонент в настоящее время не был идентифицирован, но жирные кислоты, накапливающиеся в результате разложения, были отмечены, а кусочки бумаги, покрытые синтетической олеиновой кислотой, обычно вызывали некрофоретическую реакцию. Процесс, лежащий в основе такого поведения красных огненных муравьёв, был подтвержден в 1970 году: ненасыщенные жиры, такие как олеиновая кислота, вызывают поведение по удалению трупов[34]. Рабочие также по-разному реагируют на мёртвых рабочих и куколок. Мёртвых рабочих обычно забирают из гнезда, тогда как у куколок может пройти день, чтобы возникла некрофоретическая реакция. Куколки, инфицированные Metarhizium anisopliae, обычно чаще выбрасываются рабочими; в течение суток выбрасывается 47,5 % непоражённых трупов, а у больных трупов этот показатель составляет 73,8 %[35].
Фуражировка и коммуникации
Колонии красных огненных муравьёв имеют поверхности туннелей, которые выступают за пределы поверхностей, на которых рабочие кормятся[36][37]. Эти области выступа, как правило, находятся на их собственной территории, но на это может повлиять более высокая колонизация муравьями[38]. Туннели предназначены для обеспечения эффективного взаимодействия тела, конечностей и антенн со стенами, и рабочий также может двигаться внутри них исключительно быстро (более девяти длин тела в секунду)[39]. Отверстия выходят из любой точки на территории колонии, и рабочим, собирающим пищу, может потребоваться пройти полметра, чтобы добраться до поверхности. Если предположить, что фуражир в среднем проходит 5 м, более 90 % времени сбора пищи он находится в туннелях днём и редко ночью. Рабочие собирают корм при температуре почвы, достигающей 27 °C, и температуре поверхности 12-51 °C[37]. Рабочие, подвергающиеся воздействию температуры 42 °C, рискуют умереть от жары[37]. К осени количество рабочих, собирающих пищу, быстро падает, и они редко появляются зимой. Это может быть связано с влиянием температуры почвы и снижением предпочтения источников пищи. Эти предпочтения уменьшаются только тогда, когда производство расплода низкое. В северных регионах США слишком холодно, чтобы муравьи могли добывать пищу, но в других районах, таких как Флорида и Техас, добыча пищи может происходить круглый год. Когда идёт дождь, рабочие не собирают корм на улице, так как выходные отверстия временно заблокированы, следы феромонов смываются, и дождь может физически поразить фуражиров. Влажность почвы также может влиять на поведение рабочих при поиске пищи[37]. При переноске жидкой пищи могут использовать частички почвы[40].
Когда рабочие собирают пищу, для фуражировки характерны три этапа: поиск, мобилизация и транспортировка[41]. Рабочие, как правило, ищут мёд чаще, чем другие источники пищи, и вес пищи не влияет на время поиска. Рабочие могут мобилизовать других соплеменников по гнезду, если найденная ими пища слишком тяжёлая, и для прибытия максимального количества нанятых рабочих требуется до 30 минут. Более легкие источники пищи занимают меньше времени и обычно быстро транспортируются[41]. Рабочие-фуражиры становятся разведчиками и ищут пищу за пределами поверхности и впоследствии могут умереть через две недели от старости[42].
Питание
Красные огненные муравьи — это всеядные насекомые, а фуражиры считаются падальщиками, а не хищниками[43]. Диета муравьев состоит из мертвых млекопитающих, членистоногих[44], насекомых, дождевых червей, позвоночных и твердые пищевые объекты, такие как семена. Тем не менее, этот вид предпочитает жидкие корма по сравнению с твердой пищей. Жидкая пища, которую собирают муравьи, — это сладкие вещества из растений или медвяная падь — производимая полужесткокрылыми[6][43][45][46]. Среди добываемых жертв-членистоногих могут быть двукрылые (имаго, личинки и куколок) и термиты. Известно, что потребление подслащённых аминокислот влияет на привлечение рабочих к нектарам растений. Мимические растения с сахаром редко имеют рабочих, питающимися ими, тогда как растения с сахаром и аминокислотами имеют значительное количество муравьёв[47][48]. Места обитания, в которых они живут, могут определять пищу, которую они собирают больше всего. Например, показатели успешности фуражировки для твердой пищи наиболее высоки в прибрежных районах, в то время как высокие уровни жидких источников были собраны с пастбищных участков[49]. Определенные диеты могут также влиять на рост колонии, при этом лабораторные колонии показывают высокий рост, если питаются медовой водой. Колонии, которые питаются насекомыми и сахарной водой, могут стать исключительно большими за короткий период времени, тогда как колонии, которые не питаются сахарной водой, растут значительно медленнее. Колонии, которые не питаются насекомыми, полностью прекращают выводить потомство[46]. В целом, объем пищи, перевариваемой гнездами, регулируется внутри колоний[50]. Личинки способны проявлять независимую склонность к источникам, таким как твердые белки, растворы аминокислот и растворы сахарозы, и они также предпочитают эти источники по сравнению с разбавленными растворами. Такое поведение обусловлено их способностью сообщать о голоде рабочим-нянькам. Уровень потребления зависит от типа, концентрации и состояния пищи, которой они питаются. Рабочие, как правило, привлекают больше соплеменников к источникам пищи, наполненным высоким содержанием сахарозы, чем к белковым кормам[50].
Хищники
Ряд насекомых, паукообразных и птиц охотятся на этих муравьёв, особенно когда молодые матки пытаются основать новую колонию[51]. В отсутствие рабочих матки огненных муравьёв могут полагаться на свой яд, чтобы защититься от хищников[22]. Многие виды стрекоз, в том числе Anax junius, Pachydiplax longipennis, Somatochlora provocans и Tramea carolina, захватывают маток в полёте; 16 видов пауков, в том числе паук-волк Lycosa timuga и паук чёрная вдова (Latrodectus mactans), активно убивают красных огненных муравьёв. L. mactans в своих сетях ловит все касты вида (рабочих, королев и самцов). Эти муравьи составляют 75 % добычи, пойманной пауком. Молодые пауки L. mactans также были замечены в ловле муравьев[51][52]. Среди других беспозвоночных, которые охотятся на красных огненных муравьёв отмечены уховертки (Labidura riparia) и жуки-скакуны (Cicindela punctulata)[51]. Птицы, которые поедают этих муравьёв: стриж дымчатый иглохвост (Chaetura pelagica), королевский тиранн (Tyrannus tyrannus) и виргинская американская куропатка (Colinus virginianus virginianus). Последний вид нападает на этих муравьёв, раскапывая муравейники в поисках молодых маток[51]. Красные огненные муравьи были обнаружены в содержимом желудка броненосцев[53].
Паразиты, патогены и вирусы

Известно, что мухи рода Pseudacteon (мухи-горбатки) паразитируют на муравьях. Некоторые виды этого рода, такие как Pseudacteon tricuspis, были интродуцированы в окружающую среду с целью борьбы с завезёнными огненными муравьями. Эти мухи являются паразитоидами красного огненного муравья в его естественном ареале в Южной Америке, и они привлекаются алкалоидами яда муравьёв[54]. Один вид, Pseudacteon obtusus, нападает на муравья, приземляясь на заднюю часть головы и откладывая яйцо. Расположение яйца не позволяет муравью успешно удалить его[55]. Личинки мигрируют в голову, затем развиваются в ней, питаясь гемолимфой, мышечной и нервной тканью. Примерно через две недели они заставляют голову муравья отваливаться, высвобождая фермент, растворяющий мембрану, прикрепляющую голову к его телу. Муха окукливается в отслоившейся головной капсуле и появляется через две недели[55][56]. P. tricuspis — ещё одна муха-горбатка, паразитоидная для этого вида. Хотя давление паразитизма со стороны этих мух не влияет на плотность популяции и активность муравьев, оно оказывает небольшое влияние на популяцию колонии[57]. Известно, что веерокрылое насекомое Caenocholax fenyesi (Myrmecolacidae) заражает яйца и самцов муравьёв этого вида[58][59], а клещ Pyemotes tritici (Trombidiformes) считается потенциальным биологическим агентом против красных огненных муравьёв, способных паразитировать в каждой касте внутри колония[60]. Красного огненного муравья заражают три различных варианта бактерий рода вольбахии. Однако его влияние на муравьёв неизвестно[61][62]. Solenopsis daguerrei является репродуктивным социальным паразитом в колониях красных огненных муравьёв[63].
Жизненный цикл
Брачный лёт у красных огненных муравьёв начинается в тёплое время года (весной и летом), обычно через два дня после дождя. Время появления крылатых и спаривания — между полуднем и 15:00[64][65]. Брачные полёты, зарегистрированные в Северной Флориде, включают в среднем 690 крылатых самок и самцов, участвующих в одном полете[66]. Самцы первыми покидают гнездо, и оба пола отправляются в полет практически без предполетной активности. Тем не менее, рабочие активно бегают на вершине муравейника, возбужденные феромонами максиллярных желёз крылатых половых особей[67][68]. Поскольку в холмиках муравейников нет выходов, рабочие проделывают отверстия во время брачного полета, чтобы крылья могли появиться. Это поведение рабочих, вызванное феромонами, включает быстрый бег и возвратно-поступательные движения, а также повышенную агрессию. Рабочие также группируются вокруг крылатых, когда они взбираются на растительность, и в некоторых случаях пытаются стащить их обратно, прежде чем они улетят. Химические сигналы от самцов и самок во время брачного полета привлекают рабочих, но химические сигналы, испускаемые рабочими, не привлекают других товарищей по гнезду. Это также вызывает у рабочих тревожное рекрутинговое поведение, что приводит к более высокой скорости восстановления дыхания[67][69].
Самцы летают на большей высоте, чем самки: пойманные самцы обычно находятся на высоте от 100 до 300 м над поверхностью, тогда как самки — всего от 60 до 120 м над поверхностью. Брачный полет длится примерно полчаса, и самки обычно пролетают менее 1,6 км перед приземлением. Около 95 % маток успешно спариваются и делают это только один раз; некоторые самцы могут быть бесплодны из-за того, что тестикулы не развиваются[64][70][71][72][73][74]. В полигинных колониях самцы не играют значительной роли, и поэтому большинство из них стерильны. Это также делает смертность самцов избирательной, что может повлиять на систему размножения, успех спаривания и поток генов[75][76]. Идеальные условия для начала брачного полета — это когда уровень влажности выше 80 % и температура почвы выше 18 °C. Брачные полеты происходят только при температуре окружающей среды 24-32 °C[64].
Молодых муравьиных маток часто можно найти в 2-3 км от гнезда, из которого они вылетели. Основание колонии может осуществляться индивидуально или в группах, что известно как плеометроз[77]. Эти совместные усилия маток-соосновательниц способствуют росту и выживанию зарождающейся колонии; гнезда, основанные несколькими матками, начинают период роста с в три раза большим количеством рабочих по сравнению с колониями, основанными одной маткой. Несмотря на это, такие ассоциации не всегда стабильны[78][79][80][81]. Появление первых рабочих провоцирует борьбу маток с другими матками и рабочими. В плеометротических условиях только одна королева выходит победительницей, тогда как проигравшие королевы впоследствии убиваются рабочими[82]. Два фактора, которые могут повлиять на выживание отдельных маток, — это их относительные боевые способности и их относительный вклад в производство рабочих. Размер, показатель боеспособности, положительно коррелирует с выживаемостью. Однако манипулирование относительным вкладом матки в производство рабочих не коррелировало с выживаемостью[83].
Одна матка откладывает от 10 до 15 яиц через 24 часа после спаривания[6]. В основанных развитых гнёздах матка наносит яд на каждое яйцо, которое, возможно, содержит сигнал, призывающий рабочих переместить его[84]. Эти яйца остаются неизменными в размере в течение одной недели, пока из них не вылупятся личинки. К этому времени матка отложит ещё от 75 до 125 яиц. Личинки, вылупившиеся из яиц, обычно в течение нескольких дней остаются покрытыми скорлупой. Личинки могут освобождать свой ротовой аппарат от оболочки с помощью движений тела, но всё же нуждаются в помощи рабочих при вылуплении. Личиночная стадия делится на четыре возрастные стадии, что видно по линьке. В конце каждой линьки видна часть неизвестного материала, связанная с экзувием, если они изолированы от рабочих. Личиночная стадия длится от шести до 12 дней, прежде чем их тела значительно расширятся и станут куколками; стадия куколки длится от 9 до 16 дней[6][23].
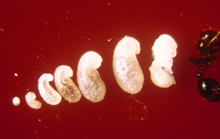
Как только первые особи достигают стадии куколки, матка прекращает яйцекладку до тех пор, пока не созреют первые рабочие. Этот процесс занимает от двух недель до одного месяца. Молодых личинок она кормит жидкостями, которые отрыгиваются из её зоба, а также трофическими яйцами или выделениями. Она также кормит расплод за счёт распада мышц крыльев, обеспечивая их необходимыми питательными веществами. Первое поколение рабочих всегда мало из-за ограниченности питательных веществ, необходимых для развития. Эти рабочие известны как минимы или нанитики, которые выкапывают выход из камеры королевы и начинают добывать пищу, необходимую для колонии. Строительство холмиков также происходит в это время. В течение месяца после рождения первого поколения начинают развиваться более крупные рабочие (солдаты), а в течение шести месяцев холмик муравейника будет заметен при осмотре и вмещает несколько тысяч обитателей. Зрелая матка способна откладывать 1500 яиц в день; все рабочие стерильны, поэтому не могут размножаться[6][85][86][87].
Колония может расти исключительно быстро. Гнёзда, в которых в мае проживало 15-20 рабочих, к сентябрю выросли до более чем 7000 муравьёв. Эти колонии начали производить репродуктивных половых особей, когда им исполнился год, и к тому времени, когда им исполнилось два года, в них насчитывалось более 25 000 рабочих. Население удвоилось до 50 000 особей, когда этим колониям исполнилось три года[88]. По достижении зрелости колония может содержать от 100 000 до 250 000 особей, но другие расчёты предполагают, что колонии могут содержать более 400 000 особей[a][89][90][91][92]. Полигинные колонии могут вырасти намного больше, чем колонии моногинные[90].
Несколько факторов способствуют росту колонии. Температура играет важную роль в росте и развитии колонии; рост колоний прекращается при температуре ниже 24°С, а время развития сокращается с 55 дней при температуре 24°С до 23 дней при 35°С. Рост развитых колоний происходит только при температуре от 24 до 36 °C. Самый первый нанитовый расплод также развивается намного быстрее, чем вторичный расплод рабочих (примерно на 35 % быстрее), что даёт преимущество для основания колоний[93]. Колонии, имеющие доступ к неограниченному количеству добываемых насекомых, существенно разрастаются, но этот рост ещё больше ускоряется, если они имеют доступ к растительным ресурсам, колонизированным полужесткокрылыми насекомыми[94]. В зарождающихся моногинных колониях, где образуются диплоидные самцы, уровень смертности колонии значительно высок, а рост колонии медленный. В некоторых случаях моногинные колонии имеют 100 % смертность на ранних стадиях развития[95].

Продолжительность жизни рабочего муравья зависит от его размера, хотя в среднем она составляет около 62 дней[96]. Мелкие рабочие живут от 30 до 60 дней, тогда как более крупные рабочие живут намного дольше. Более крупные рабочие, продолжительность жизни которых составляет от 60 до 180 дней, живут на 50-140 % дольше, чем их более мелкие собратья, но это зависит от температуры[6][97]. Однако известно, что рабочие, содержащиеся в лабораторных условиях, живут от 10 до 70 недель (от 70 до 490 дней); максимальная зарегистрированная продолжительность жизни рабочего составляет 97 недель (или 679 дней)[98]. Матки живут намного дольше, чем рабочие, от двух почти до семи лет[6][98].
Матки — единственные муравьи в колонии, способные изменять соотношение полов. Например, королевы, происходящие из колоний, производящих самцов, как правило, производят преимущественно самцов, в то время как королевы, происходящие из колоний с предпочтительным соотношением полов в пользу самок, как правило, производят самок[99][100]. Матки также контролируют производство половых особей с помощью феромонов, которые влияют на поведение рабочих по отношению к личинкам как самцов, так и самок[101].
Моногинные и полигинные формы
У огненных муравьёв встречаются две формы сообществ: полигинные и моногинные колонии[102][103]. У общественных насекомых полигинные колонии существенно отличаются от моногинных, как по числу маток, так и по степени успешности в расселении, продолжительности жизни маток и в значении коэффициента родственности соплеменников по гнезду[104][105][106][107]. Полигинные матки также менее физогастричны, чем моногинные матки, а их рабочие мельче[108][109]. Количество старых маток в колонии не влияет на набор или привлечение новых самок (в будущем новых маток) в семью. Важную роль в регуляции размножения играют концентрации феромонов маток, которые, по-видимому, связаны с числом маток. Из этого следует, что рабочие отказываются от привлечения новых самок (маток), когда подвергаются воздействию большого количества этого маточного феромона. Более того, полученные доказательства подтверждают утверждение о том, что самки в обеих популяциях входят в гнезда наугад, без какого-либо отношения к числу обитающих там старых королев[110]. При этом нет никакой корреляции между числом старых маток и числом вновь принятых в семью молодых самок. Были высказаны три гипотезы, объясняющие принятие множества самок в сложившиеся развитые колонии: мутуализм, родственный отбор (kin selection) и паразитизм[111]. В гипотезе мутуализма утверждается, что сотрудничество приводит к увеличению личной приспособленности старших маток. Однако эта гипотеза не согласуется с тем, что увеличение числа королев уменьшает как продуктивность маток, так и их долговечность[112]. Родственный отбор (kin selection) также кажется маловероятным, учитывая, что матки, как было обнаружено, кооперируют в условиях, когда они статистически не родственны[113]. Таким образом, матки не получают никакой выгоды в личной приспособленности, при пополнении колонии новыми самками. Лучшим объяснением полигинии уже существующих гнезд представляется паразитизм. Одно из предположений заключается в том, что многие самки пытаются войти в колонию, чтобы рабочие запутались и невольно позволили нескольким самкам присоединиться к ней[113].
Моногинные рабочие убивают чужих самок и агрессивно защищают свою территорию. Однако не все поведения универсальны, прежде всего потому, что поведение рабочих зависит от экологического контекста, в котором они развиваются, а манипуляции с генотипами рабочих могут вызывать изменения в поведении. Поэтому поведение местных популяций может отличаться от поведения внедренных популяций[114].
При исследовании агрессивного поведения моногинных и полигинных огненных муравьёв обнаружено, что этограммы представителей разного типа колоний легко отличаются[115]. Рабочие из моногинных колоний всегда нападали на вторгающихся муравьев из чужих колоний независимо от того, были ли они из моногинных или полигиновых колоний, тогда как рабочие из полигинных колоний опознавали, но не нападали на чужих муравьёв из полигинных колоний, главным образом, демонстрируя предупреждающие позы, подобные поведению, принятому после атак паразитических мух Phseudacteon. Предупреждающее поведение сильно зависело от социальной структуры рабочих. Поэтому поведение по отношению к чужим рабочим служит методом характеризации моногинных и полигинных колоний[116]. Большинство колоний на юго-востоке и южном центре США имеют тенденцию быть моногинными[81].
Размер территории и величина муравейника у моногинных колоний красного огненного муравья положительно коррелированы, что, в свою очередь, регулируется размером колонии (численность и биомасса рабочих), расстоянием от соседних колоний, плотностью добычи и коллективной конкурентоспособностью колонии. Напротив, дискриминация соплеменников среди полигинных колоний значительно ослаблена, поскольку рабочие терпят конспецифических муравьёв, чуждых их колонии, принимают других самок и не агрессивно защищают свою территорию от вторгшихся рабочих из других полигинных колоний[117]. Полигинные колонии увеличивают их репродуктивные способности в результате того, что у них много маток, больше возможности использования крупных территорий посредством кооперативного мобилизации из многих взаимосвязанных муравейников[118].
Влияние на другие виды организмов

Когда полигинные формы вторгаются в новые места обитания, то разнообразие местных членистоногих и позвоночных снижается[119]. Значительно снижается численность популяций изопод, клещей и Scarabaeidae. Они также могут значительно изменить численность популяций представителей многих семейств мух и жуков, включая: Calliphoridae, Histeridae, Muscidae, Sarcophagidae, Silphidae и Staphylinidae. Несмотря на это общее явление, было обнаружено, что некоторые виды насекомых могут не пострадать от красных огненных муравьев. Например, если плотность изопод уменьшается в очагах заражения красными огненными муравьями, то сверчки рода Gryllus не подвержены такому влиянию. Известны случаи, когда разнообразие некоторых видов насекомых и членистоногих увеличивается в районах, где присутствуют красные огненные муравьи[120][121][122]. Красные огненные муравьи являются важными хищниками пещерных беспозвоночных, некоторые из которых находятся под угрозой исчезновения. Среди них сенокосцы, ложноскорпионы, пауки, наземные жуки и Pselaphinae. Самую большую опасность представляет не сам муравей, а ядовитые приманки, используемые против него. Считается, что хищнические нападения красных огненных муравьев является основным фактором исчезновения древесной улитки Orthalicus reses, которые вымерли в дикой природе[120]. В целом, красные огненные муравьи предпочитают конкретных членистоногих другим, хотя они атакуют и убивают любого беспозвоночного, который не может защитить себя или убежать[123]. Биоразнообразие членистоногих возрастает, как только популяции красных импортированных муравьев сокращаются или уничтожаются[10][124].
Взаимодействие красных огненных муравьев и млекопитающих редко регистрируется. Тем не менее, отмечены случаи смерти животных, попавших в ловушки и подвергшихся нападениям красных огненных муравьев[120][125][126]. В результате взаимодействия между огненными муравьями и флоридским кроликом (Sylvilagus floridanus), показатели смертность молоди этих млекопитающих возрастают от 33 до 75 %[127]. Обнаружено, что красные огненные муравьи оказывают сильное влияние на многие виды герпетофауны. Наблюдается снижение численности населения обыкновенной королевской змеи (Lampropeltis getula floridana), а также яиц и взрослых особей заборной игуаны (Sceloporus undulatus) и американских варанов (Aspidoscelis sexlineata), используемых муравьями в качестве источник пищи[128][129][130]. Естественный отбор, вызванный нападениями красных огненных муравьев, привел к тому, что некоторые ящерицы (например, Sceloporus undulatus) стали иметь более длинные ноги и новое поведение, чтобы избежать их атак[131][132]. Взрослые коробчатые черепахи (Terrapene carolina triunguis), молодые хьюстонские жабы (Anaxyrus houstonensis) и только что вылупившиеся миссисипские аллигаторы (Alligator mississippiensis) также атакуются и убиваются красными огненными муравьями[120][133][134][135]. Несмотря на эту, в основном, отрицательную ассоциацию, обнаружено, что красные импортированные муравьи могут оказывать влияние на трансмиссивные болезни путем регулирования популяций клещей и изменения динамики вектора и хозяина, тем самым снижая скорость передачи не только на животных, но и на людей[136].
Птицы
В результате нападений этих муравьёв у некоторых птиц возрастают показатели смертности. Были случаи, когда в районах с высокой плотностью муравьев молодые птенцы вообще не доживали до взрослой жизни. Многие птицы, в том числе ласточки, утки, цапли, перепела и крачки подвергаются воздействию красных огненных муравьёв[137]. Особенно уязвимы для нападений огненных муравьёв гнездящиеся на земле птицы, например, американские крачки (Sterna antillarum)[138][135]. Влияние красных огненных муравьёв на колониально гнездящихся птиц очень велико, смертность околоводных птиц из-за их нападений может достигать 100 %, хотя этот фактор был ниже у рано гнездящихся птиц. Смертность молоди у белолобой горной ласточки (Petrochelidon pyrrhonota) возрастает от нападения фуражирующих рабочих муравьёв[139]. Певчие воробьиные также демонстрируют увеличение смертности в областях распространения красных огненных муравьёв, а уровни выживаемости Vireo griseus (Vireo griseus) и Vireo atricapilla (Vireo atricapilla) возрастают от 10 % до 31 % и от 7 % до 13 %, соответственно, в тех местах где огненные муравьи отсутствуют, или они стали неспособны атаковать птиц[140].
Красные огненные муравьи также оказывают косвенное влияние на выживание молоди лугового тетерева (Tympanuchus cupido attwateri)[141]. Сначала считалось, что муравьи непосредственно связаны со снижением численности перезимовавших птиц, таких как американский жулан (Lanius ludovicianus), но более поздние исследования показали, что усилия по искоренению муравьев с использованием пестицида Mirex, который, как известно, имеет токсические побочные эффекты, в значительной степени имеют основное воздействие на снижение численности птиц[142][143].
Взаимодействие с другими видами муравьёв
Красные огненные муравьи являются сильными конкурентами для многих видов муравьёв. Им удалось вытеснить многие коренные виды муравьёв, что привело к ряду экологических последствий[144]. Однако, исследования показывают, что эти муравьи не всегда превосходят конкурентов и не всегда подавляют коренных муравьёв. Более правдоподобными причинами нарушения численности коренных муравьёв считается нарушение условий их обитания и местной экологии ещё до прибытия импортированных видов[145]. Красный огненный муравей сильнее видов Tapinoma melanocephalum и Pheidole fervida, но показывает различные уровни агрессии по отношению к ним. Например, они менее враждебны к T. melanocephalum, чем к P. fervida. Уровень смертности у T. melanocephalum и P. fervida при столкновении с красными огненными муравьями разнятся и составляют 31,8 % и 49,9 %, соответственно. При этом уровень смертности самих огненных муравьёв составляет только от 0,2 % до 12 %[146]. Муравей Рэсберри (Nylanderia fulva) демонстрирует большее доминирование, чем красный огненный муравей, и даже вытесняет их в местах обитания, где они сталкиваются друг с другом[147]. Большие колонии дерновых муравьёв (Tetramorium caespitum) могут уничтожить семьи красных огненных муравьёв, что позволяет энтомологам сделать вывод о том, что этот конфликт между двумя видами может помочь помешать распространению красного огненного муравья[148][149]. Муравьи, инфицированные SINV-1 погибают быстрее, чем здоровые индивидуумы муравьёв Monomorium chinense. Это означает, что муравьи, заражённые SINV-1 слабее, чем их здоровые соплеменники и быстрее элиминируются муравьями M. chinense. Однако, крупные рабочие, заражённые или нет, погибают редко[150].
В районах, где он является нативным, красный огненный муравей по-прежнему оказывается доминирующим видом и сосуществует с 28 видами муравьёв в галерейных лесах и с 10 видами в ксерофитных лесных лугах, выигрывая в агрессивных взаимодействиях с другими муравьями. Однако, некоторые муравьи могут быть содоминантными в областях, где они сосуществуют, например, аргентинский муравей конкурирует симметрично[151][152]. Рабочие огненных муравьёв регулярно участвуют в конкурентных взаимоотношениях за кормовые ресурсы с другими муравьями и могут подавлять использование пищевых ресурсов, например, от сосущих соки растений полужесткокрылых насекомых (в частности, кокцид вида Phenacoccus solenopsis) от местных муравьёв. Однако, красные огненные муравьи не могут устранить T. melanocephalum полностью, хотя, они в итоге потребляют более высокую долю пищи. Вместо этого два муравья могут мирно сосуществовать и делиться кормовым ресурсом[153][154]. Когда они встречаются с огненными муравьями, рабочие могут демонстрировать поведение видимой смерти и притворяются мёртвыми, что позволяет им успешно избегать опасность. Однако такое поведение наблюдается только у молодых рабочих, поскольку старые рабочие либо убегают, либо сопротивляются, когда им угрожают[155].
Красные огненные муравьи также конкурируют с аргентинскими муравьями. Показатели смертности варьируются в разных сценариях (например, смертность в конфронтации колоний ниже, чем у тех, кто индивидуально сталкивается друг с другом в этой области). Крупные рабочие (солдаты) также могут выдержать больше травм на своих телах, тем самым увеличивая смертность аргентинских муравьёв. Для аргентинских муравьёев, чтобы успешно уничтожить моногинную колонию огненных муравьёв в 160 000 рабочих, потребуется своя колонии численностью 396 800 особей. Колония, которая уменьшилась в размерах благодаря успешной обработке ядовитой приманкой, подвержена хищничеству аргентинских муравьёв. Аргентинские муравьи могут играть жизненно важную роль в удалении ослабленных колоний огненных муравьёв, и они также могут быть важны в замедлении распространения этих муравьёв, особенно в областях, сильно заражённых аргентинскими муравьями[156]. Несмотря на это, популяции аргентинских муравьёв в юго-восточной части Соединённых Штатов сократились после появления красного огненного муравья[157].

Красные огненные муравьи образуют взаимовыгодные симбиотические отношения с инвазивными кокцидами Phenacoccus solenopsis. Рост колоний увеличивается, если муравьи имеют доступ к ресурсам, созданным Ph. solenopsis, а плотность популяции этих мучнистых мух значительно выше в районах присутствия красных огненных муравьёв (распространение Ph. solenopsis в районах, где эти кокциды не является родными, связано с наличием муравьёв). Кроме того, ожидаемая продолжительность жизни и репродуктивная скорость Ph. solenopsis возрастают. Эти червецы могут даже переноситься рабочими муравьями обратно в своё гнездо[158]. Поедание Ph. solenopsis их врагами уменьшается из-за того, что рабочие препятствуют хищничеству и паразитизму со стороны естественных врагов. Червецы поедаются жуками божьими коровками (Menochilus sexmaculatus) и являются хозяевами для двух паразитов (Aenasius bambawalei и Acerophagus coccois), если рядом нет муравьёв. Но если присутствуют муравьи, то божьи коровки реже встречаются на таких растениях, а скорость движения их личинок значительно ниже[158]. Это ясно видно на примере популяций хлопковой тли (Aphis gossypii) в районах с присутствием красного огненного муравья[159][160][161]. Красные огненные муравьи активно участвуют в развитии мутуалистических связей с кокцидами Dysmicoccus morrisoni. Муравьи вызывают рост популяции D. morrisoni, защищая их от врагов, покрывая колонии защитными укрытиями и собирая, выделяемые ими сладкие соки (падь)[162].
Систематика
Таксономия огненных муравьёв сложна и, кроме того, они образуют гибриды (например, гибрид красного огненного муравья с чёрным огненным муравьём (Solenopsis richteri)[163][164][165]. До 1972 года все огненные муравьи фигурировали под названием Solenopsis saevissima Smith, 1855 (однако, сам этот вид встречается только в Южной Америке). Красный огненный муравей в старой литературе обозначался в качестве красной формы его чёрного подвида Solenopsis saevissima richteri Forel, 1909. Кладистический анализ с использованием данных ДНК показал, что красный огненный муравей вместе с несколькими родственными видами (Solenopsis saevissima Smith, 1855, Solenopsis daguerrei Santschi, 1930, S. richteri, и другими) относится к группе видов Solenopsis saevissima species-group. К этой группе также относится комплекс близких видов Solenopsis geminata complex (Solenopsis aurea, S. geminata, S. xyloni и другие виды)[166].
Филогения
Красный огненный муравей включён в состав видовой группы S. saevissima species-group. Филогенетические отношения красного огненного муравья с близкими видами этой группы и с S. geminata отображены на кладограмме[167][168][169]:
| Solenopsis |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Значение

Яд огненных муравьёв содержит алкалоид соленопсин (транс-2-метил-6-n-ундецилпиперидин) и оказывает дерматонекротическое, цитотоксическое, гемолитическое, нейротоксическое и инсектицидное действие. Степень воздействия по индексу силы ужалений Шмидта оценивается в 1,2 балла и сходна с воздействием огня (отсюда и название насекомого). У людей, ужаленных этими муравьями, часто наблюдается аллергическая реакция, покраснение кожи, отёк, волдыри, тошнота, рвота, головокружения и реакции вплоть до анафилактического шока (отмечены случаи со смертельным исходом)[170]. Особенную опасность укусы огненных муравьёв представляют для детей[171].
Общий ущерб в США составляет около 5 млрд долларов ежегодно, с учётом затрат на медицинское обслуживание ужаленных муравьями, ветеринарный контроль домашних животных, уничтожение муравейников и химобработку территорий[172]. Тем не менее влияние огненных муравьев на природу неоднозначно: являясь агрессивными хищниками, они регулируют популяции многих вредителей сельского хозяйства, в том числе вредителей сахарного тростника, хлопчатника, риса и зерновых культур. Например, они сокращают численность таких вредителей как хлопковый долгоносик (Curculionidae)[173][174][175], огнёвки-травянки Diatraea saccharalis (Crambidae) на полях сахарного тростника[176][177], мухи Haematobia irritans (Diptera: Muscidae)[178], совки Anticarsia gemmatalis на соевых бобах, белокрылки в оранжереях[179][180].
Укусы и ужаления огненных муравьёв негативно воздействуют на домашних животных и представителей дикой фауны, в том числе изменяют поведение и морфологию ящериц[181]. Также красные огненные муравьи могут вытеснять местные виды муравьёв. Например, в южных штатах США они почти вытеснили своих родственников, местных огненных муравьёв — Solenopsis xyloni и Solenopsis geminata[182].
Способы борьбы
Биологические методы
Для борьбы с огненными муравьями используют следующие организмы: микроспоридии вида Thelohania solenopsae, грибы Myrmicinosporidium durum[183] и Beauveria bassiana, социально-паразитического муравья Solenopsis daguerrei, мух-горбаток (Phoridae; виды Pseudacteon tricuspis и Pseudacteon curvatus)[184], другие организмы[185].
Физические методы
Для истребления красных огненных муравьёв первоначально проводилась выкопка гнёзд. Однако матки могли уходить по подземным ходам на глубину более 1 метра, что приводило к скорому возобновлению колонии. Затем стали использоваться кипящая вода и даже жидкий азот, которые оказались эффективными в истреблении муравейников[186].
Химические методы
Первоначально в США применялись пестициды марки «Мирекс», которые распыляли с вертолётов. Но в 1976 году было доказано, что эти вещества опасны для птиц и человека и являются сердечным токсином[187].
Генетика
Геном: 0,62 пг (C value)[188][189]. Диплоидный набор хромосом 2n = 32[190].
Исследования показывают, что вариации митохондриальной ДНК в значительной степени встречаются в полигинных семьях (гнёзда с несколькими матками)[191], но никаких изменений не обнаруживается в моногинных сообществах (гнёзда с одной маткой)[192]. Триплоидия (хромосомная аномалия) часто встречается у красных огненных муравьев (до 12% у нерепродуктивных самок), что связано с высокой частотой диплоидных самцов[193]. Красный огненный муравей - первый вид, обладающий геном green-beard gene, благодаря которому естественный отбор может способствовать альтруистическому поведению. Рабочие, у которых есть этот ген, могут различать маток, содержащих его, и маток, у которых нет, по-видимому, с помощью запаховых сигналов. Рабочие убивают маток, не содержащих гена[194][195]. В 2011 году ученые объявили, что полностью секвенировали геном красного огненного муравья[196].
См. также
Примечания
Комментарии
Источники
- ↑ Малышева М. С. Муравьи // Насекомые и клещи ― вредители сельскохозяйственных культур / Под редакцией Нарчук Э. П., Тряпицын В. А.. — Л.: Наука, 1981. — Т. IV. Перепончатокрылые и двукрылые. — С. 48. — 222 с.
- ↑ Феоктистова Н. Ю. Огненные муравьи–переселенцы. Еженедельник «Биология. Первое сентября», № 24 (2003). Дата обращения: 3 августа 2015. Архивировано 20 августа 2016 года.
- ↑ Константин Болотов. Горбатая муха должна обезглавить огненного муравья. Мембрана (22 декабря 2005). Дата обращения: 3 августа 2015. Архивировано 5 марта 2016 года.
- ↑ Ольга Кувыкина. Муравьи — людоеды. Вокруг света (июль 2015). Дата обращения: 3 августа 2015. Архивировано 6 мая 2015 года.
- ↑ Стриганова Б. Р., Захаров А. А. Пятиязычный словарь названий животных: Насекомые. Латинский, русский, английский, немецкий, французский / под ред. д-ра биол. наук, проф. Б. Р. Стригановой. — М.: РУССО, 2000. — С. 295. — 1060 экз. — ISBN 5-88721-162-8.
- ↑ 1 2 3 4 5 6 7 8 Laura Collins, Rudolf H. Scheffrahn. Red imported fire ant (англ.). UF/IFAS Featured Creatures. University of Florida (декабрь 2016). Дата обращения: 20 июня 2017. Архивировано 21 сентября 2016 года.
- ↑ Red Imported Fire Ant (англ.). National Invasive Species Information Center. USDA. Дата обращения: 20 июня 2017. Архивировано 12 июля 2017 года.
- ↑ Ahmedin Jemal, Martin Hugh-Jones. A review of the red imported fire ant (Solenopsis invicta Buren) and its impacts on plant, animal, and human health (англ.) // Preventive Veterinary Medicine. — 1993. — October (vol. 17, no. 1—2). — P. 19—32. — doi:10.1016/0167-5877(93)90051-T.
- ↑ 1 2 3 4 Wetterer, James Kelly. Exotic spread of Solenopsis invicta Buren (Hymenoptera: Formicidae) beyond North America (англ.) // Sociobiology. — Feira de Santana, BA, Brazil, 2013. — Vol. 60, no. 1. — P. 50—55. — ISSN 2447-8067. Архивировано 4 июля 2017 года.
- ↑ 1 2 Epperson D.M., Allen C.R. Red imported fire ant impacts on upland arthropods in southern Mississippi (англ.) // The American Midland Naturalist : журнал. — 2010. — Vol. 163, no. 1. — P. 54—63. — doi:10.1674/0003-0031-163.1.54.
- ↑ Callcott A.-M., Collins H.L. Invasion and range expansion of imported fire ants (Hymenoptera: Formicidae) in North America from 1918–1995 (англ.) // The Florida Entomologist : журнал. — 1996. — Vol. 79. — P. 240—251. — doi:10.2307/3495821. — .
- ↑ Korzukhin M.D., Porter S.D., Thompson L.C., Wiley S. Modeling temperature-dependent range limits for the fire ant Solenopsis invicta (Hymenoptera: Formicidae) in the United States (англ.) // Environmental Entomology : журнал. — 2001. — Vol. 30, no. 4. — P. 645—655. — doi:10.1603/0046-225X-30.4.645. Архивировано 4 марта 2016 года.
- ↑ McCubbin K., Weiner J. Fire ants in Australia: a new medical and ecological hazard (англ.) // Medical Journal of Australia. — 2002. — Vol. 176, no. 11. — P. 518—9. — PMID 12064981.
- ↑ Огненный подарок на Лунный Новый год Архивная копия от 20 октября 2013 на Wayback Machine.
- ↑ Список 80 ДИССЕРТАЦИЙ об огненных муравьях Архивная копия от 24 мая 2012 на Wayback Machine.
- ↑ USDA Quarantine Map. Дата обращения: 2 мая 2010. Архивировано 9 июня 2012 года.
- ↑ Hedges, S.A. Handbook of Pest Control. — 8th. — Mallis Handbook and Technical Training Company, 1997. — P. 531–535.
- ↑ 1 2 3 4 5 6 7 8 9 Buren W. F. Revisionary studies on the taxonomy of the imported fire ants (англ.) // Journal of the Georgia Entomological Society : журнал. — 1972. — Vol. 7. — P. 1—26. — doi:10.5281/zenodo.27055. Архивировано 9 апреля 2016 года.
- ↑ Kevin L. Haight. 2010. Worker Size and Nest Defense in Solenopsis invicta (Hymenoptera: Formicidae) Архивная копия от 17 марта 2022 на Wayback MachineAnnals of the Entomological Society of America. 103(4):678—682. 2010. (англ.) (Дата обращения: 28 ноября 2010)
- ↑ Santschi, F. (1916). "Formicides sudaméricains nouveaux ou peu connus". Physis. 2: 365—399. doi:10.5281/ZENODO.14374.
- ↑ Walter, David ; Moser, John. (2010). Gaeolaelaps Invictianus, a new and unusual species of Hypoaspidine Mite (Acari: Mesostigmata: Laelapidae) phoretic on the red imported fire ant Solenopsis Invicta Buren (Hymenoptera: Formicidae) in Louisiana, USA Архивная копия от 15 октября 2011 на Wayback Machine. International Journal of Acarology 36(5):399-407
- ↑ 1 2 Fox, Eduardo G.P.; Wu, Xiaoqing; Wang, Lei; Chen, Li; Lu, Yong-Yue; Xu, Yijuan (February 2019). "Queen venom isosolenopsin A delivers rapid incapacitation of fire ant competitors". Toxicon. 158: 77—83. doi:10.1016/j.toxicon.2018.11.428. PMID 30529381.
- ↑ 1 2 3 Petralia, R. S.; Vinson, S. B. (1979). "Developmental morphology of larvae and eggs of the Imported fire ant, Solenopsis invicta". Annals of the Entomological Society of America. 72 (4): 472—484. doi:10.1093/aesa/72.4.472.
- ↑ 1 2 Wheeler G. C., Wheeler J. (1977). "Supplementary studies on ant larvae: Myrmicinae". Transactions of the American Entomological Society. 103: 581—602. doi:10.5281/zenodo.25100.
{{cite journal}}: Википедия:Обслуживание CS1 (множественные имена: authors list) (ссылка) - ↑ Petralia, R.S.; Sorensen, A.A.; Vinson, S.B. (1980). "The labial gland system of larvae of the imported fire ant, Solenopsis invicta Buren". Cell and Tissue Research. 206 (1): 145—156. doi:10.1007/BF00233615. PMID 6153574.
- ↑ Greenberg, L.; Kabashima, J.N. Pest Notes: Red Imported Fire Ant. Statewide Integrated Pest Management Program. Agriculture and Natural Resources, University of California (2014). Дата обращения: 4 апреля 2016. Архивировано 21 мая 2016 года.
- ↑ O'Neal, J.; Markin, G.P. (1975). "The larval instars of the imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae)". Journal of the Kansas Entomological Society. 48 (2): 141—151. JSTOR 25082731.
- ↑ 1 2 3 4 Fox, Eduardo G P; Solis, Daniel Russ; Rossi, Mônica Lanzoni; Delabie, Jacques Hubert Charles; de Souza, Rodrigo Fernando; Bueno, Odair Correa (2012). "Comparative Immature Morphology of Brazilian Fire Ants (Hymenoptera: Formicidae: Solenopsis)". Psyche: A Journal of Entomology. 2012: 1—10. doi:10.1155/2012/183284.
- ↑ Mlot, N.J.; Craig, A.T.; Hu, D.L. (2011). "Fire ants self-assemble into waterproof rafts to survive floods". Proceedings of the National Academy of Sciences. 108 (19): 7669—7673. Bibcode:2011PNAS..108.7669M. doi:10.1073/pnas.1016658108. PMC 3093451. PMID 21518911.
- ↑ Morrill, W.L. (1974). "Dispersal of red imported fire ants by water". The Florida Entomologist. 57 (1): 39—42. doi:10.2307/3493830. JSTOR 3493830.
- ↑ Hölldobler & Wilson, 1990, p. 171.
- ↑ 1 2 Adams, B.J.; Hooper-Bùi, L.M.; Strecker, R.M.; O'Brien, D.M. (2011). "Raft formation by the red imported fire ant, Solenopsis invicta". Journal of Insect Science. 11 (171): 171. doi:10.1673/031.011.17101. PMC 3462402. PMID 22950473.
- ↑ Haight, K.L. (2006). "Defensiveness of the fire ant, Solenopsis invicta, is increased during colony rafting". Insectes Sociaux. 53 (1): 32—36. doi:10.1007/s00040-005-0832-y.
- ↑ Howard, D.F.; Tschinkel, W.R. (1976). "Aspects of necrophoric behavior in the red imported fire ant, Solenopsis invicta" (PDF). Behaviour. 56 (1): 157—178. doi:10.1163/156853976x00334. ISSN 0005-7959. Архивировано (PDF) из оригинала 23 февраля 2022. Дата обращения: 23 февраля 2022.
- ↑ Qiu, H.-L.; Lu, L.-H.; Shi, Q.-X.; Tu, C.-C.; Lin, T.; He, Y.-R. (2015). "Differential necrophoric behaviour of the ant Solenopsis invicta towards fungal-infected corpses of workers and pupae". Bulletin of Entomological Research. 105 (5): 607—614. doi:10.1017/S0007485315000528. PMID 26082426.
- ↑ Markin G.P., O'Neal J., Dillier J. Foraging tunnels of the red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae) (англ.) // Journal of the Kansas Entomological Society. — 1975. — Vol. 48, no. 1. — P. 83–89. — .
- ↑ 1 2 3 4 Porter S.D., Tschinkel W.R. Foraging in Solenopsis invicta (Hymenoptera: Formicidae): effects of weather and season (англ.) // Environmental Entomology. — 1987. — Vol. 16, no. 3. — P. 802–808. — doi:10.1093/ee/16.3.802.
- ↑ Showler A.T., Knaus R.M., Reagan T.E. Foraging territoriality of the imported fire ant, Solenopsis invicta Buren, in surgarcane as determined by neutron activation analysis (англ.) // Insectes Sociaux. — 1989. — Vol. 36, no. 3. — P. 235–239. — doi:10.1007/BF02226306.
- ↑ Gravish N., Monaenkova D., Goodisman M.A.D., Goldman D.I. Climbing, falling, and jamming during ant locomotion in confined environments (англ.) // Proceedings of the National Academy of Sciences. — 2013. — Vol. 110, no. 24. — P. 9746–9751. — doi:10.1073/pnas.1302428110. — . — arXiv:1305.5860. — PMID 23690589. — PMC 3683784.
- ↑ Qin W, Chen X, Hooper-Bùi LM, Cai J, Wang L, Sun Z, Wen X, Wang C. Food-burying behavior in red imported fire ants (Hymenoptera: Formicidae) (англ.) // PeerJ. — 2019. — Vol. 7, no. e6349. — doi:10.7717/peerj.6349. Архивировано 30 января 2019 года.
- ↑ 1 2 Yijuan X., Yongyue L., Ling Z., Guangwen L. Foraging behavior and recruitment of red imported fire ant Solenopsis invicta Buren in typical habitats of South China (англ.) // Acta Ecologica Sinica. — 2007. — Vol. 27, no. 3. — P. 855–860. — doi:10.1016/S1872-2032(07)60022-5.
- ↑ Tschinkel W.R. The organization of foraging in the fire ant, Solenopsis invicta (англ.) // Journal of Insect Science. — 2011. — Vol. 11, no. 26. — P. 26. — doi:10.1673/031.011.0126. — PMID 21529150. — PMC 3391925.
- ↑ 1 2 Taber, 2000, p. 37.
- ↑ Taber, 2000, p. 38.
- ↑ Tennant, L.E.; Porter, S.D. Comparison of diets of two fire ant species (Hymenoptera: Formicidae): solid and liquid components (англ.) // Journal of Entomological Science : журнал. — 1991. — Vol. 26, no. 4. — P. 450—465. — ISSN 0749-8004.
- ↑ 1 2 Porter, S.D. Effects of diet on the growth of laboratory fire ant colonies (Hymenoptera: Formicidae) (англ.) // Journal of the Kansas Entomological Society : журнал. — 1989. — Vol. 62, no. 2. — P. 288—291. — .
- ↑ Lanza, J.; Vargo, E. L.; Pulim, S.; Chang, Y. Z. Preferences of the fire ants Solenopsis invicta and S. geminata (Hymenoptera: Formicidae) for amino acid and sugar components of extrafloral nectars (англ.) // Environmental Entomology : журнал. — 1993. — Vol. 22, no. 2. — P. 411—417. — doi:10.1093/ee/22.2.411.
- ↑ Lanza, J. Response of fire ants (Formicidae: Solenopsis invicta and S.gerninata) to artificial nectars with amino acids (англ.) // Ecological Entomology : журнал. — 1991. — Vol. 16, no. 2. — P. 203—210. — doi:10.1111/j.1365-2311.1991.tb00210.x.
- ↑ Vogt, J.T.; Grantham, R.A.; Corbett, E.; Rice, S.A.; Wright, R.E. Dietary habits of Solenopsis invicta (Hymenoptera: Formicidae) in four Oklahoma habitats (англ.) // Environmental Entomology : журнал. — 2002. — Vol. 31, no. 1. — P. 47—53. — doi:10.1603/0046-225X-31.1.47.
- ↑ 1 2 Cassill, D.L.; Tschinkel, W.R. Regulation of diet in the fire ant, Solenopsis invicta (англ.) // Journal of Insect Behavior : журнал. — 1999. — Vol. 12, no. 3. — P. 307—328. — doi:10.1023/A:1020835304713. Архивировано 26 января 2021 года.
- ↑ 1 2 3 4 Whitcomb, W.H.; Bhatkar, A.; Nickerson, J.C. (1973). "Predators of Solenopsis invicta queens prior to successful colony establishment". Environmental Entomology. 2 (6): 1101—1103. doi:10.1093/ee/2.6.1101.
- ↑ Nyffeler, M.; Dean, D.A.; Sterling, W.L. (1988). "The southern black widow spider, Latrodectus mactans (Araneae, Theridiidae), as a predator of the red imported fire ant, Solenopsis invicta (Hymenoptera, Formicidae), in Texas cotton fields". Journal of Applied Entomology. 106 (1—5): 52—57. doi:10.1111/j.1439-0418.1988.tb00563.x.
- ↑ Deyrup, M.; Trager, J. (1986). "Ants of the Archbold Biological Station, Highlands County, Florida (Hymenoptera: Formicidae)". The Florida Entomologist. 69 (1): 206—228. doi:10.2307/3494763. JSTOR 3494763. Архивировано из оригинала 24 февраля 2020. Дата обращения: 17 марта 2021.
- ↑ Chen, Li; Sharma, K.R.; Fadamiro, H.Y. (2009). "Fire ant venom alkaloids act as key attractants for the parasitic phorid fly, Pseudacteon tricuspis (Diptera: Phoridae)". Naturwissenschaften. 96 (12): 1421—1429. Bibcode:2009NW.....96.1421C. doi:10.1007/s00114-009-0598-6. PMID 19714317.
- ↑ 1 2 Williams, D.F.; Banks, W.A. (1987). "Pseudacteon obtusus (Diptera: Phoridae) attacking Solenopsis invicta (Hymenoptera: Formicidae) in Brazil". Psyche: A Journal of Entomology. 94 (1—2): 9—13. doi:10.1155/1987/85157.
- ↑ Ehrenberg, R. (2009-09-18). "Venom attracts decapitating flies". Science News. Архивировано из оригинала 22 сентября 2016. Дата обращения: 30 декабря 2014. (требуется подписка)
- ↑ Morrison, L.W.; Porter, S.D. (2005). "Testing for population-level impacts of introduced Pseudacteon tricuspis flies, phorid parasitoids of Solenopsis invicta fire ants". Biological Control. 33 (1): 9—19. CiteSeerX 10.1.1.542.4377. doi:10.1016/j.biocontrol.2005.01.004.
- ↑ Schmid-Hempel, 1998, p. 94.
- ↑ Kathirithamby, J.; Johnston, J.S. (1992). "Stylopization of Solenopsis invicta (Hymenoptera: Formicidae) by Caenocholax fenyesi (Strepsiptera: Myrmecolacidae) in Texas". Annals of the Entomological Society of America. 85 (3): 293—297. doi:10.1093/aesa/85.3.293.
- ↑ Bruce, W.A.; LeCato, G.L. (1980). "Pyemotes tritici: A potential new agent for biological control of the red imported fire ant, Solenopsis invicta (Acari: Pyemotidae)". International Journal of Acarology. 6 (4): 271—274. doi:10.1080/01647958008683230.
- ↑ Ahrens, M.E.; Shoemaker, D. (2005). "Evolutionary history of Wolbachia infections in the fire ant Solenopsis invicta". BMC Evolutionary Biology. 5: 35. doi:10.1186/1471-2148-5-35. PMC 1175846. PMID 15927071.
{{cite journal}}: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (ссылка) - ↑ Schmid-Hempel, 1998, p. 34.
- ↑ Capinera, 2008, p. 188.
- ↑ 1 2 3 Trager, 1988, p. 419.
- ↑ Buhs, 2005, p. 21.
- ↑ Morrill, W.L. (1974). "Production and flight of alate red imported fire ants". Environmental Entomology. 3 (2): 265—271. doi:10.1093/ee/3.2.265.
- ↑ 1 2 Alonso, L.E.; Vander Meer, R.K. (1997). "Source of alate excitant pheromones in the red imported fire ant Solenopsis invicta (Hymenoptera: Formicidae)". Journal of Insect Behavior. 10 (4): 541—555. doi:10.1007/BF02765376.
- ↑ Markin, G.P.; Dillier, J.H.; Hill, S.O.; Blum, M.S.; Hermann, H.R. (1971). "Nuptial flight and flight ranges of the imported fire ant, Solenopsis saevissima richteri (Hymenoptera: Formicidae)". Journal of the Georgia Entomological Society. 6 (3): 145—156.
- ↑ Obin, M.S.; Vander Meer, R.K. (1994). "Alate semiochemicals release worker behavior during fire ant nuptial flights" (PDF). Journal of Entomological Science. 29 (1): 143—151. doi:10.18474/0749-8004-29.1.143. Архивировано (PDF) из оригинала 20 августа 2016. Дата обращения: 23 февраля 2022.
- ↑ Hung, A.C.F.; Vinson, S. B.; Summerlin, J. W. (1974). "Male sterility in the red imported fire ant, Solenopsis invicta". Annals of the Entomological Society of America. 67 (6): 909—912. doi:10.1093/aesa/67.6.909.
- ↑ Trager, 1988, p. 422.
- ↑ Capinera, 2008, p. 3444.
- ↑ Capinera, 2008, p. 3112.
- ↑ Hölldobler & Wilson, 1990, p. 156.
- ↑ Fritz, G.N.; Vander Meer, R.K.; Preston, C.A. (2006). "Selective male mortality in the red imported fire ant, Solenopsis invicta". Genetics. 173 (1): 207—213. doi:10.1534/genetics.106.056358. PMC 1461442. PMID 16489215.
- ↑ Hölldobler & Wilson, 1990, p. 145.
- ↑ Tschinkel, W. (1992). "Brood raiding and the population dynamics of founding and incipient colonies of the fire ant, Solenopsis invicta". Ecological Entomology. 17 (2): 179—188. doi:10.1111/j.1365-2311.1992.tb01176.x.
- ↑ Goodisman, M.; Ross, K. (1999). "Queen recruitment in a multiple-queen population of the fire ant Solenopsis invicta". Behavioral Ecology. 10 (4): 428—435. doi:10.1093/beheco/10.4.428.
- ↑ Tschinkel, W.R.; Howard, D.F. (1983). "Colony founding by pleometrosis in the fire ant, Solenopsis invicta" (PDF). Behavioral Ecology and Sociobiology. 12 (2): 103—113. doi:10.1007/BF00343200. Архивировано (PDF) из оригинала 23 февраля 2022. Дата обращения: 23 февраля 2022.
- ↑ Trager, 1988, p. 217.
- ↑ 1 2 Trager, 1988, p. 218.
- ↑ Manfredini, F.; Riba-Grognuz, O.; Wurm, Y.; Keller, L.; Shoemaker, D.; Grozinger, C.M.; Tsutsui, N.D. (2013). "Sociogenomics of cooperation and conflict during colony founding in the fire ant Solenopsis invicta". PLOS Genetics. 9 (8): e1003633. doi:10.1371/journal.pgen.1003633. PMC 3738511. PMID 23950725.
{{cite journal}}: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (ссылка) - ↑ Bernasconi, G.; Keller, L. (1996). "Reproductive conflicts in cooperative associations of fire ant queens (Solenopsis invicta)" (PDF). Proceedings of the Royal Society B: Biological Sciences. 263 (1369): 509—513. Bibcode:1996RSPSB.263..509B. doi:10.1098/rspb.1996.0077. Архивировано (PDF) из оригинала 12 сентября 2021. Дата обращения: 23 февраля 2022.
- ↑ Taber, 2000, p. 42.
- ↑ Doebeli, Mi.; Abouheif, E. (2016). "Modeling evolutionary transitions in social insects". eLife. 5: e12721. doi:10.7554/eLife.12721. PMC 4744194. PMID 26780668.
{{cite journal}}: Википедия:Обслуживание CS1 (не помеченный открытым DOI) (ссылка) - ↑ Hölldobler & Wilson, 1990, p. 157.
- ↑ Hölldobler & Wilson, 1990, p. 168.
- ↑ Markin, G. P.; Dillier, J. H.; Collins, H. L. (1973). "Growth and development of colonies of the red imported fire ant, Solenopsis invicta". Annals of the Entomological Society of America. 66 (4): 803—808. doi:10.1093/aesa/66.4.803.
- ↑ 1 2 Hölldobler & Wilson, 1990, p. 163.
- ↑ 1 2 Taber, 2000, p. 32.
- ↑ Schmid-Hempel, 1998, p. 10.
- ↑ Schmid-Hempel, 1998, p. 16.
- ↑ Porter, S.D. (1988). "Impact of temperature on colony growth and developmental rates of the ant, Solenopsis invicta". Journal of Insect Physiology. 34 (12): 1127—1133. doi:10.1016/0022-1910(88)90215-6.
- ↑ Helms, K.R.; Vinson, S.B. (2008). "Plant resources and colony growth in an invasive ant: the importance of honeydew-producing Hemiptera in carbohydrate transfer across trophic levels". Environmental Entomology. 37 (2): 487—493. doi:10.1093/ee/37.2.487. PMID 18419921.
- ↑ Ross, K.G.; Fletcher, D.J.C. (1986). "Diploid male production — a significant colony mortality factor in the fire ant Solenopsis invicta (Hymenoptera: Formicidae)". Behavioral Ecology and Sociobiology. 19 (4): 283—291. doi:10.1007/BF00300643.
- ↑ Wang, L.; Xu, Y.; Di, Z.; Roehner, B.M. (2013). "How does group interaction and its severance affect life expectancy?" (PDF). Cornell University Library: 1—28. arXiv:1304.2935. Bibcode:2013arXiv1304.2935W. S2CID 14848627. Архивировано из оригинала (PDF) 3 ноября 2016.
- ↑ Calabi, P.; Porter, S.D. (1989). "Worker longevity in the fire ant Solenopsis invicta: Ergonomic considerations of correlations between temperature, size and metabolic rates". Journal of Insect Physiology. 35 (8): 643—649. doi:10.1016/0022-1910(89)90127-3.
- ↑ 1 2 Hölldobler & Wilson, 1990, p. 170.
- ↑ Davies, N.B. An Introduction to Behavioural Wcology / N.B. Davies, J.R. Krebs, S.A. West. — 4th. — Chichester : Wiley-Blackwell, 2012. — P. 385. — ISBN 978-1-4051-1416-5.
- ↑ Passera, L.; Aron, S.; Vargo, E.L.; Keller, L. (2001). "Queen control of sex Ratio in fire ants" (PDF). Science. 293 (5533): 1308—1310. Bibcode:2001Sci...293.1308P. doi:10.1126/science.1062076. PMID 11509728. Архивировано (PDF) из оригинала 23 февраля 2022. Дата обращения: 23 февраля 2022.
- ↑ Vargo, E.L.; Fletcher, D.J.C. (1986). "Evidence of pheromonal queen control over the production of male and female sexuals in the fire ant, Solenopsis invicta". Journal of Comparative Physiology A. 159 (6): 741—749. doi:10.1007/BF00603727. ISSN 1432-1351.
- ↑ Tschinkel, 2006, p. 95.
- ↑ Hölldobler & Wilson, 1990, p. 187.
- ↑ Goodisman Michael A., D., Ross K.G. A test of queen recruitment models using nuclear and mitochondrial markers in the fire ant Solenopsis invicta (англ.) // Evolution : журнал. — Wiley-VCH, 1998. — Vol. 52, no. 5. — P. 1416. — doi:10.2307/2411311. — . Архивировано 5 мая 2015 года.
- ↑ Morel L., Meer R.K.V., Lofgren C.S. Comparison of nestmate recognition between monogyne and polygyne populations of Solenopsis invicta (Hymenoptera: Formicidae) (англ.) // Annals of the Entomological Society of America : журнал. — 1990. — Vol. 83, no. 3. — P. 642—647. — doi:10.1093/aesa/83.3.642.
- ↑ Schmid-Hempel, 1998, p. 191.
- ↑ Trager, 1988, p. 204.
- ↑ Fletcher D. J. C., Blum M. S., Whitt T. V., Temple N. Monogyny and polygyny in the fire ant, Solenopsis invicta (англ.) // Annals of the Entomological Society of America : журнал. — 1980. — Vol. 73, no. 6. — P. 658—661. — doi:10.1093/aesa/73.6.658.
- ↑ Greenberg L., Fletcher D.J.C., Vinson S.B. Differences in worker size and mound distribution in monogynous and polygynous colonies of the fire ant Solenopsis invicta Buren (англ.) // Journal of the Kansas Entomological Society : журнал. — 1985. — Vol. 58, no. 1. — P. 9—18. — .
- ↑ Harrison, R.G. Dispersal polymorphisms in insects (неопр.) // Annual Review of Ecology and Systematics. — 1980. — Т. 11, № 1. — С. 95—118. — doi:10.1146/annurev.es.11.110180.000523. — .
- ↑ DeHeer C.J., Goodisman M.A.D., Ross K.G. Queen dispersal strategies in the multiple‐queen form of the fire ant Solenopsis invicta (англ.) // The American Naturalist : журнал. — University of Chicago Press, 1999. — Vol. 153, no. 6. — P. 660—675. — ISSN 0003-0147. — doi:10.1086/303205. — . Архивировано 24 сентября 2015 года.
- ↑ Keller, L. Social life: the paradox of multiple-queen colonies (англ.) // Trends in Ecology & Evolution : журнал. — 1995. — Vol. 10, no. 9. — P. 355—360. — doi:10.1016/S0169-5347(00)89133-8. — PMID 21237068.
- ↑ 1 2 Ross, Kenneth G. The breeding system of the fire ant Solenopsis invicta: Effects on colony genetic structure (англ.) // The American Naturalist : журнал. — University of Chicago Press, 1993. — Vol. 141, no. 4. — P. 554—576. — doi:10.1086/285491. — PMID 19425999. — .
- ↑ Goodisman, M.A.D.; Sankovich, K.A.; Kovacs, J.L. Genetic and morphological variation over space and time in the invasive fire ant Solenopsis invicta (англ.) // Biological Invasions : журнал. — 2006. — Vol. 9, no. 5. — P. 571—584. — doi:10.1007/s10530-006-9059-8. Архивировано 24 сентября 2015 года.
- ↑ Adams, E.S.; Balas, M.T. Worker discrimination among queens in newly founded colonies of the fire ant Solenopsis invicta (англ.) // Behavioral Ecology and Sociobiology : журнал. — 1999. — Vol. 45, no. 5. — P. 330—338. — doi:10.1007/s002650050568.
- ↑ Chirino, M.G.; Gilbert, L.E.; Folgarait, P.J. Behavioral discrimination between monogyne and polygyne red fire ants (Hymenoptera: Formicidae) in their native range (англ.) // Annals of the Entomological Society of America : журнал. — 2012. — Vol. 105, no. 5. — P. 740—745. — doi:10.1603/AN11073.
- ↑ Goodisman, M.; Ross, K.G. Relationship of queen number and queen relatedness in multiple-queen colonies of the fire ant Solenopsis invicta (англ.) // Ecological Entomology : журнал. — 1997. — Vol. 22, no. 2. — P. 150—157. — doi:10.1046/j.1365-2311.1997.t01-1-00052.x.
- ↑ Adams, E.S.; Tschinkel, W.R. Mechanisms of population regulation in the fire ant Solenopsis invicta: an experimental study (англ.) // Journal of Animal Ecology : журнал. — 2001. — Vol. 70, no. 3. — P. 355—369. — doi:10.1046/j.1365-2656.2001.00501.x.
- ↑ The reduction in the biodiversity of Australian native fauna and flora due to the red imported fire ant, Solenopsis invicta (fire ant). Department of the Environment and Energy. Government of Australia. Дата обращения: 10 августа 2016. Архивировано 22 сентября 2016 года.
- ↑ 1 2 3 4 Wojcik, D.; Allen, C. R.; Brenner, R. J.; Forys, E.A.; Jouvenaz, D. P.; Lutz, R.S. Red imported fire ants: impact on biodiversity (неопр.) // American Entomologist. — 2001. — Т. 47, № 1. — С. 16—23. — doi:10.1093/ae/47.1.16. Архивировано 16 декабря 2010 года.
- ↑ Porter, S.D.; Savignano, D.A. Invasion of polygyne fire ants decimates native ants and disrupts arthropod community (англ.) // Ecology : журнал. — 1990. — Vol. 71, no. 6. — P. 2095—2106. — doi:10.2307/1938623.
- ↑ Stoker, R. L.; Grant, W.E.; Bradleigh Vinson, S. Solenopsis invicta (Hymenoptera: Formicidae) effect on invertebrate decomposers of carrion in central Texas (англ.) // Environmental Entomology : журнал. — 1995. — Vol. 24, no. 4. — P. 817—822. — doi:10.1093/ee/24.4.817.
- ↑ Ricks, B.L.; Vinson, B.S. Feeding acceptability of certain insects and various water-soluble compounds to two varieties of the imported fire ant (англ.) // Journal of Economic Entomology : журнал. — 1970. — Vol. 63, no. 1. — P. 145—148. — doi:10.1093/jee/63.1.145.
- ↑ Cook, J.L. Conservation of biodiversity in an area impacted by the red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae) (англ.) // Biodiversity and Conservation : журнал. — 2003. — Vol. 12, no. 2. — P. 187—195. — doi:10.1023/A:1021986927560.
- ↑ Masser, M.P.; Grant, W.E. Fire ant-Induced trap mortality of small mammals in east-central Texas (англ.) // The Southwestern Naturalist : журнал. — 1986. — Vol. 31, no. 4. — P. 540—542. — doi:10.2307/3671712. — .
- ↑ Flickinger, E.L. Observations of predation by red imported fire ants on live-trapped wild cotton rats (англ.) // Texas Journal of Science : журнал. — 1989. — Vol. 41, no. 2. — P. 223—224.
- ↑ Hill, E.P. Observations of imported fire ant predation on nestling cottontails (англ.) // Proceedings of the Annual Conference, Southeastern Association of Game and Fish Commissioners : журнал. — 1970. — Vol. 23. — P. 171—181.
- ↑ Thawley, C.J.; Langkilde, T. Invasive fire ant (Solenopsis invicta) predation of eastern fence lizard (Sceloporus undulatus) eggs (англ.) // Journal of Herpetology : журнал. — 2016. — Vol. 50, no. 2. — P. 284—288. — doi:10.1670/15-017.
- ↑ Mount, R.H.; Trauth, S.E.; Mason, W.H. Predation by the red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae), on eggs of the lizard Cnemidophorus sexlineatus (Squamata: Teiidae) (англ.) // Journal of the Alabama Academy of Science : журнал. — 1981. — Vol. 52. — P. 66—70.
- ↑ Bartlett, D. 40 years of thoughts on Paynes Prairie (неопр.) // Reptiles. — 1997. — Т. 5, № 7. — С. 68, 70—73.
- ↑ Langkilde, T. Invasive fire ants alter behavior and morphology of native lizards (англ.) // Ecology : журнал. — 2009. — Vol. 90, no. 1. — P. 208—217. — doi:10.1890/08-0355.1.
- ↑ Bryner, J. (2009-01-26). "Lizards' Dance Avoids Deadly Ants". LiveScience. Архивировано из оригинала 4 марта 2016. Дата обращения: 20 августа 2016.
- ↑ Freed, P.S.; Neitman, K. Notes on predation on the endangered Houston toad, Bufo houstonensis (англ.) // Texas Journal of Science : журнал. — 1988. — Vol. 40. — P. 454—456.
- ↑ Allen, C.R.; Rice, K.G.; Wojcik, D.P.; Percival, H.F. Effect of red imported fire ant envenomization on neonatal American alligators (англ.) // Journal of Herpetology : журнал. — 1997. — Vol. 31, no. 2. — P. 318—321. — doi:10.2307/1565408.
- ↑ 1 2 Capinera, 2008, p. 1080.
- ↑ Castellanos Adrian A., Medeiros Matthew C. I., Hamer Gabriel L., Morrow Michael E.; Eubanks, Micky D.; Teel, Pete D.; Hamer, Sarah A.; Light, Jessica E. Decreased small mammal and on-host tick abundance in association with invasive red imported fire ants (Solenopsis invicta) (англ.) // Biology Letters : журнал. — Royal Society Publishing, 2016. — Vol. 12, no. 9. — P. 20160463. — doi:10.1098/rsbl.2016.0463. — PMID 27651533. — PMC 5046925.
- ↑ Moloney, S.D.; Vanderwoude, C. Potential ecological impacts of red imported fire ants in eastern Australia (англ.) // Journal of Agricultural and Urban Entomology : журнал. — 2003. — Vol. 20, no. 3. — P. 131—142. Архивировано 9 октября 2016 года.
- ↑ Lockley, T.C. Effect of imported fire ant predation on a population of the least tern: an endangered species (англ.) // Southwestern Entomologist : журнал. — 1995. — Vol. 20. — P. 517—519.
- ↑ Sikes, P.J.; Arnold, K.A. Red imported fire ant (Solenopsis invicta) predation on cliff swallow (Hirundo pyrrhonota) nestlings in east central Texas (англ.) // Southwestern Naturalist : журнал. — 1986. — Vol. 31. — P. 105—106. — doi:10.2307/3670967.
- ↑ Campomizzi, A.J.; Morrison, M.L.; Farrell, S.L.; Wilkins, R..N.; Drees, B.M.; Packard, J.M. Red imported fire ants can decrease songbird nest survival (англ.) // The Condor : журнал. — 2009. — Vol. 111, no. 3. — P. 534—537. — doi:10.1525/cond.2009.090066.
- ↑ Morrow, M.E.; Chester, R.E.; Lehnen, S.E.; Drees, B.M.; Toepfer, J.E. Indirect effects of red imported fire ants on Attwater's prairie-chicken brood survival (англ.) // The Journal of Wildlife Management : журнал. — 2015. — Vol. 79, no. 6. — P. 898—906. — doi:10.1002/jwmg.915. Архивировано 22 августа 2016 года.
- ↑ Yosef, R.; Lohrer, F.E. Loggerhead shrikes, red fire ants and red herrings? (англ.) // The Condor : журнал. — 1995. — Vol. 97, no. 4. — P. 1053—1056. — doi:10.2307/1369544. — . Архивировано 13 августа 2017 года.
- ↑ "Environment: Fighting the Fire Ant". Time. 1970-11-02. Архивировано из оригинала 22 сентября 2016. Дата обращения: 28 августа 2016.
- ↑ Capinera, 2008, p. 1771.
- ↑ King J.R., Tschinkel W.R. Experimental evidence that the introduced fire ant, Solenopsis invicta, does not competitively suppress co-occurring ants in a disturbed habitat (англ.) // Journal of Animal Ecology : журнал. — 2006. — Vol. 75, no. 6. — P. 1370—1378. — doi:10.1111/j.1365-2656.2006.01161.x. — PMID 17032369. — .
- ↑ Gao, Y.; Lu, L-H.; He, Y-R.; Qi, G-J.; Zhang, J-Q. Interference competition between the red imported fire ant (Solenopsis invicta Buren) and two native ant species (Hymenoptera: Formicidae) (англ.) // Acta Entomologica Sinica : журнал. — 2011. — Vol. 54, no. 5. — P. 602—608.
- ↑ LeBrun, E.G.; Abbott, J.; Gilbert, L.E. Imported crazy ant displaces imported fire ant, reduces and homogenizes grassland ant and arthropod assemblages (англ.) // Biological Invasions : журнал. — 2013. — Vol. 15, no. 11. — P. 2429—2442. — doi:10.1007/s10530-013-0463-6.
- ↑ Vitone T., Lucky A. Pavement ant - Tetramorium caespitum. UF/IFAS Featured Creatures. University of Florida (2014). Дата обращения: 4 апреля 2016. Архивировано 19 апреля 2016 года.
- ↑ King, T.G.; Phillips, S.A. Destruction of young colonies of the red imported fire ant by the pavement ant (англ.) // Entomological News : журнал. — 1992. — Vol. 103, no. 3. — P. 72—77. — ISSN 0013-872X.
- ↑ Chen, Y.C.; Kafle, L.; Shih, C.J. Interspecific competition between Solenopsis invicta and two native ant species, Pheidole fervens and Monomorium chinense (англ.) // Journal of Economic Entomology : журнал. — 2011. — Vol. 104, no. 2. — P. 614—621. — doi:10.1603/ec10240. — PMID 21510213.
- ↑ Calcaterra, L.A.; Livore, J.P.; Delgado, A.; Briano, J.A. Ecological dominance of the red imported fire ant, Solenopsis invicta, in its native range (англ.) // Oecologia : журнал. — 2008. — Vol. 156, no. 2. — P. 411—421. — doi:10.1007/s00442-008-0997-y. (недоступная ссылка)
- ↑ LeBrun E.G., Tillberg C.V., Suarez A.V., Folgarait P.J., Smith C.R.; Holway, D.A. An experimental study of competition between fire ants and Argentine ants in their native range. (англ.) // Ecology : журнал. — 2007. — Vol. 88, no. 1. — P. 63—75. — doi:10.1890/0012-9658(2007)88[63:AESOCB]2.0.CO;2. — PMID 17489455.
- ↑ Zhou A., Liang G., Lu Y., Zeng L., Xu Y. Interspecific competition between the red imported fire ant, Solenopsis invicta Buren and ghost ant, Tapinoma melanocephalum Fabricius for honeydew resources produced by an invasive mealybug, Phenacoccus solenopsis Tinsiley (англ.) // Arthropod-Plant Interactions : журнал. — 2014. — Vol. 8, no. 5. — P. 469—474. — doi:10.1007/s11829-014-9324-1.
- ↑ Wu B., Wang L., Liang G., Lu Y.; Zeng, L. Food competition mechanism between Solenopsis invicta Buren and Tapinoma melanocephalum Fabricius (англ.) // Sociobiology : журнал. — 2014. — Vol. 61, no. 3. — P. 265—273. — doi:10.13102/sociobiology.v61i3.265-273. Архивировано 22 сентября 2016 года. Архивированная копия. Дата обращения: 12 июля 2017. Архивировано 22 сентября 2016 года.
- ↑ Cassill D.L., Vo K., Becker B. Young fire ant workers feign death and survive aggressive neighbors (англ.) // Naturwissenschaften : журнал. — 2008. — Vol. 95, no. 7. — P. 617—624. — doi:10.1007/s00114-008-0362-3. — . — PMID 18392601.
- ↑ Kabashima J.N., Greenberg L.; Rust, M.K.; Paine, T.D. Aggressive interactions between Solenopsis invicta and Linepithema humile (Hymenoptera: Formicidae) under laboratory conditions. (англ.) // Journal of Economic Entomology : журнал. — 2007. — Vol. 100, no. 1. — P. 148—154. — doi:10.1603/0022-0493(2007)100[148:AIBSIA]2.0.CO;2. — PMID 17370822. Архивировано 9 октября 2016 года.
- ↑ Capinera, 2008, p. 290.
- ↑ 1 2 Zhou A., Lu Y., Zeng L.; Xu, Y.; Liang, G. Does mutualism drive the invasion of two alien species? The case of Solenopsis invicta and Phenacoccus solenopsis (англ.) // PloS ONE : журнал. — 2012. — Vol. 7, no. 7. — P. e41856. — doi:10.1371/journal.pone.0041856. — . — PMID 22911859. — PMC 3402455.
- ↑ Kaplan I., Eubanks M.D. Disruption of cotton aphid (Homoptera: Aphididae)—natural enemy dynamics by red imported fire ants (Hymenoptera: Formicidae) (англ.) // Environmental Entomology : журнал. — 2002. — Vol. 31, no. 6. — P. 1175—1183. — doi:10.1603/0046-225X-31.6.1175.
- ↑ Coppler L.B., MurphyJ.F., Eubanks M.D. Red imported fire ants (Hymenoptera: Formicidae) increase the abundance of aphids in tomato (англ.) // The Florida Entomologist : журнал. — 2007. — Vol. 90, no. 3. — P. 419—425. — doi:10.1653/0015-4040(2007)90[419:RIFAHF]2.0.CO;2.
- ↑ Diaz R., Knutson A., Bernal J.S. Effect of the red imported fire ant on cotton aphid population density and predation of bollworm and beet armyworm eggs. (англ.) // Journal of Economic Entomology : журнал. — 2004. — Vol. 97, no. 2. — P. 222—229. — doi:10.1603/0022-0493-97.2.222. — PMID 15154439.
- ↑ Tedders, W.L.; Reilly, C.C.; Wood, B.W.; Morrison, R.K.; Lofgren, C.S. Behavior of Solenopsis invicta (Hymenoptera: Formicidae) in pecan orchards (англ.) // Environmental Entomology : журнал. — 1990. — Vol. 19, no. 1. — P. 44—53. — doi:10.1093/ee/19.1.44.
- ↑ Vander Meer, R.K., Lofgren, C.S. & Alvarez, F.M. (1985) Biochemical evidence for hybridization in fire ants. Florida Entomologist, 68, 501—506.
- ↑ Ross, K.G., Vander Meer, R.K., Fletcher, D.J.C. & Vargo, E.L. (1987) Biochemical phenotypic and genetic studies of two introduced fire ant species and their hybrid (Hymenoptera: Formicidae). Evolution, 41, 280—293.
- ↑ Shoemaker, D.D., Ross, K.G. & Arnold, M.L. (1996) Genetic structure and evolution of a fire ant hybrid zone. — Evolution, 50, 1958—1976.
- ↑ Кладистический анализ огненных муравьев. Архивная копия от 20 октября 2013 на Wayback Machine «Zoologica Scripta», 2005, Volume 34, Issue 5, Pages 493—505
- ↑ Tschinkel, W. (2006). The Fire Ants. Cambridge, Massachusetts: Belknap Press of Harvard University Press. ISBN 978-0-674-02207-2.
- ↑ Pitts, J.P.; McHugh, J.V.; Ross, K.G. Cladistic analysis of the fire ants of the Solenopsis saevissima species-group (Hymenoptera: Formicidae) (англ.) // Zoologica Scripta : журнал. — 2005. — Vol. 34, no. 5. — P. 493—505. — doi:10.1111/j.1463-6409.2005.00203.x.
- ↑ Pitts, J.P. (2002). A cladistic analysis of the Solenopsis saevissima species-group (Hymenoptera: Formicidae) (PDF) (Ph.D.). University of Georgia, Athens. pp. 42—43. Архивировано (PDF) из оригинала 23 марта 2012. Дата обращения: 23 февраля 2017.
- ↑ Яд огненных муравьев Solenopsis invicta. Дата обращения: 2 мая 2010. Архивировано 20 октября 2009 года.
- ↑ Эдвард Уилсон. Планета муравьёв = Wilson Edward. Tales from the Ant World. — М.: Альпина нон-фикшн , 2022. — С. 42. — 211 с. — ISBN 978-5-00139-583-6.
- ↑ McDonald, Maggie. Reds Under Your Feet (interview with Robert Vander Meer) (неопр.) // New Scientist. — 2006. — February (т. 189, № 2538). — С. 50.
- ↑ Sterling, W.L. Fortuitous biological suppression of the boll weevil by the red imported fire ant (англ.) // Environmental Entomology : журнал. — 1978. — Vol. 7, no. 4. — P. 564—568. — doi:10.1093/ee/7.4.564.
- ↑ Fillman, D.A.; Sterling, W.L. Killing power of the red imported fire ant (Hym.: Formicidae): a key predator of the boll weevil [Col.: Curculionidae] (англ.) // Entomophaga : журнал. — 1983. — Vol. 28, no. 4. — P. 339—344. — doi:10.1007/BF02372186.
- ↑ Sterling, W.L.; Dean, D.A.; Fillman, D.A.; Jones, D. Naturally-occurring biological control of the boll weevil [Col.: Curculionidae] (англ.) // Entomophaga : журнал. — 1984. — Vol. 29, no. 1. — P. 1—9. — doi:10.1007/BF02372203.
- ↑ Negm, A.A.; Hensley, S.D. The relationship of arthropod predators to crop damage inflicted by the sugarcane borer (англ.) // Journal of Economic Entomology : журнал. — 1967. — Vol. 60, no. 6. — P. 1503—1506. — doi:10.1093/jee/60.6.1503.
- ↑ Reagan, T.E.; Coburn, G.; Hensley, S.D. Effects of mirex on the arthropod fauna of a Louisiana sugarcane field (англ.) // Environmental Entomology : журнал. — 1972. — Vol. 1, no. 5. — P. 588—591. — doi:10.1093/ee/1.5.588.
- ↑ Lemke, L.A.; Kissam, J.B. Impact of red imported fire ant (Hymenoptera: Formicidae) predation on horn flies (Diptera: Muscidae) in a cattle pasture treated with pro-drone (англ.) // Journal of Economic Entomology : журнал. — 1988. — Vol. 81, no. 3. — P. 855—858. — doi:10.1093/jee/81.3.855. — PMID 3403766.
- ↑ Lee, J.-H.; Johnson, S.J.; Wright, V.L. Quantitative survivorship analysis of the velvetbean caterpillar (Lepidoptera: Noctuidae) pupae in soybean fields in Louisiana (англ.) // Environmental Entomology : журнал. — 1990. — Vol. 19, no. 4. — P. 978—986. — doi:10.1093/ee/19.4.978.
- ↑ Morrill, W.L. Red imported fire ant foraging in a greenhouse (неопр.) // Environmental Entomology. — 1977. — Т. 6, № 3. — С. 416—418. — doi:10.1093/ee/6.3.416.
- ↑ Инвазивные муравьи изменяют поведение и морфологию ящериц Архивная копия от 3 февраля 2014 на Wayback Machine. «Ecology», 2009, Volume 90(1), pp. 208—217
- ↑ Solenopsis xyloni. (недоступная ссылка) — Invasive Ant Treat. Information Sheet Number 29. landcareresearch.co.nz.
- ↑ Всем грибам на борьбу с огненным муравьем! Дата обращения: 2 мая 2010. Архивировано 13 февраля 2012 года.
- ↑ Мухи-фориды против огненных муравьев Solenopsis invicta. Дата обращения: 2 мая 2010. Архивировано 20 октября 2013 года.
- ↑ Интернационал социальных и микропаразитов против огненных муравьев. Дата обращения: 2 мая 2010. Архивировано 20 октября 2013 года.
- ↑ Nature & Science " Biology Resources " Integrated Pest Management Manual. Дата обращения: 2 мая 2010. Архивировано 22 сентября 2016 года.
- ↑ Муравьи — людоеды Архивная копия от 31 октября 2010 на Wayback Machine в журнале «Вокруг Света» (2006)
- ↑ База данных о размерах геномов животных. Дата обращения: 16 мая 2010. Архивировано 1 февраля 2014 года.
- ↑ Li, J. and K. M. Heinz (2000). Genome complexity and organization in the red imported fire ant Solenopsis invicta Buren. Genetical Research 75: 129—135.
- ↑ Lorite P.& Palomeque T. Karyotype evolution in ants (Hymenoptera: Formicidae) with a review of the known ant chromosome numbers. Архивная копия от 7 июня 2012 на Wayback Machine — Myrmecologische Nachrichten (Wien). — 2010. Volume 13, Pages 89-102. (Дата обращения: 12 декабря 2010)
- ↑ Taber, 2000, p. 43.
- ↑ Ross, K.G.; Shoemaker, D.D. (1997). "Nuclear and mitochondrial genetic structure in two social forms of the fire ant Solenopsis invicta: insights into transitions to an alternate social organization". Heredity. 78 (6): 590—602. doi:10.1038/hdy.1997.98.
- ↑ Krieger, M.J.B.; Ross, K.G.; Chang, C..W.Y.; Keller, L. (1999). "Frequency and origin of triploidy in the fire ant Solenopsis invicta". Heredity. 82 (2): 142—150. doi:10.1038/sj.hdy.6884600.
- ↑ Keller, L.; Ross, K.G. (1998). "Selfish genes: a green beard in the red fire ant" (PDF). Nature. 394 (6693): 573—575. Bibcode:1998Natur.394..573K. doi:10.1038/29064. Архивировано (PDF) из оригинала 20 июля 2011. Дата обращения: 13 марта 2021.
- ↑ Grafen, A. (1998). "Evolutionary biology: Green beard as death warrant" (PDF). Nature. 394 (6693): 521—522. Bibcode:1998Natur.394..521G. doi:10.1038/28948. Архивировано (PDF) из оригинала 3 сентября 2021. Дата обращения: 13 марта 2021.
- ↑ Wurm, Y.; Wang, J.; Riba-Grognuz, O.; Corona, M.; Nygaard, S.; Hunt, B.G.; Ingram, K.K.; Falquet, L.; Nipitwattanaphon, M.; Gotzek, D.; Dijkstra, M.B.; Oettler, J.; Comtesse, F.; Shih, C.-J.; Wu, W.-J.; Yang, C.-C.; Thomas, J.; Beaudoing, E.; Pradervand, S.; Flegel, V.; Cook, E.D.; Fabbretti, R.; Stockinger, H.; Long, L.; Farmerie, W.G.; Oakey, J.; Boomsma, J.J.; Pamilo, P.; Yi, S.V.; Heinze, J.; Goodisman, M. A.D.; Farinelli, L.; Harshman, K.; Hulo, N.; Cerutti, L.; Xenarios, I.; Shoemaker, D.; Keller, L. (2011). "The genome of the fire ant Solenopsis invicta" (PDF). Proceedings of the National Academy of Sciences. 108 (14): 5679—5684. Bibcode:2011PNAS..108.5679W. doi:10.1073/pnas.1009690108. PMC 3078418. PMID 21282665. Архивировано (PDF) из оригинала 9 октября 2016. Дата обращения: 13 марта 2021.
Литература
- Buren W. F. Revisionary studies on the taxonomy of the imported fire ants (англ.) // Journal of the Georgia Entomological Society : журнал. — 1972. — Vol. 7. — P. 1—26. — doi:10.5281/zenodo.27055.
- Ward D. 2009: The potential distribution of the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae), in New Zealand. New Zealand entomologist, 32: 67—75.
- Buhs J.B. The Fire Ant Wars: Nature, Science and Public Policy in Twentieth Century America. — Chicago, Illinois, 2005. — ISBN 978-0-226-07981-3.
- Capinera J.L. Encyclopedia of Entomology. — 2nd. — Dordrecht, Netherlands: Springer, 2008. — ISBN 978-1-4020-6242-1.
- Hölldobler B., Wilson E.O. The Ants. — Cambridge, Massachusetts: Belknap Press of Harvard University Press, 1990. — ISBN 978-0-674-04075-5.
- Schmid-Hempel. Parasites in Social Insects. — Princeton, New Jersey: Princeton University Press, 1998. — ISBN 978-0-691-05924-2.
- Taber S.W. Fire Ants. — College Station, Texas: Texas A&M University Press, 2000. — ISBN 978-1-60344-711-9.
- Trager J.C. Advances in Myrmecology. — 1st. — Leiden, Netherlands: E.J. Brill, 1988. — ISBN 978-0-916846-38-1.
- Tschinkel W. The Fire Ants. — Cambridge, Massachusetts: Belknap Press of Harvard University Press, 2006. — ISBN 978-0-674-02207-2.
Ссылки
- Яд огненного муравья Solenopsis invicta. Дата обращения: 14 июля 2017.
- Laura Collins, Rudolf H. Scheffrahn. Red imported fire ant (англ.). UF/IFAS Featured Creatures. University of Florida (2016). Дата обращения: 14 июля 2017.
- Огненные муравьи на сайте National Pest Management Association (англ.). Дата обращения: 14 июля 2017.
- Огненные муравьи на сайте eXtension (англ.). Дата обращения: 14 июля 2017. Архивировано из оригинала 6 октября 2007 года.
- Огненные муравьи в базе данных генной информации
- Австралия
- Огненные муравьи в базе Department of Primary Industries & Fisheries, Queensland (англ.). Дата обращения: 14 июля 2017. Архивировано из оригинала 6 октября 2017 года.
- США
- Огненные муравьи в базе National Invasive Species Information Center (англ.). invasiveSpeciesinfo.gov. Agricultural Research Service USDA (2017). Дата обращения: 14 июля 2017.
- Огненные муравьи в Техасе: Texas Imported Fire Ant Research and Management Project (англ.). Дата обращения: 14 июля 2017.
- Обзор исследований в области токсикохимии яда красных огненных муравьёв Solenopsis invicta и других видов. Дата обращения: 14 июля 2017.
- Огненные муравьи на сайте Global Invasive Species Database. (англ.). Дата обращения: 14 июля 2017.
| Таксономия | |
|---|---|
| В библиографических каталогах |
Обычно почти сразу, изредка в течении часа.