

В биохимии аллостерическая регуляция (или аллостерический контроль) — это регуляция фермента путем связывания эффекторной молекулы в сайте, отличном от активного сайта фермента[1].
Сайт, с которым связывается эффектор, называется аллостерическим сайтом или регуляторным сайтом. Аллостерические сайты позволяют эффекторам связываться с белком, что часто приводит к конформационным изменениям, связанным с динамикой белка. Эффекторы, которые усиливают активность белка, называются аллостерическими активаторами, тогда как те, которые снижают активность белка, называются аллостерическими ингибиторами.
Аллостерические регуляции — естественный пример контуров управления, таких как обратная связь от последующих продуктов или прямая связь от исходных субстратов. Аллостерия дальнего действия особенно важна для передачи сигналов клетками[2]. Аллостерическая регуляция также особенно важна для способности клетки регулировать активность ферментов.
Термин аллостерия происходит от древнегреческого allos (ἄλλος), «прочее» и stereos (στερεὀς), «твердое тело (объект)». Это относится к тому факту, что регуляторный сайт аллостерического белка физически отличается от его активного сайта.
Модели
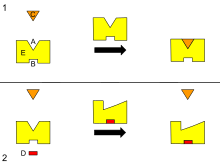
B — Аллостерический сайт
C — Субстрат
D — Ингибитор
E — Фермент
Диаграмма аллостерической регуляции фермента.
Многие аллостерические эффекты могут быть объяснены согласованной моделью MWC выдвинутых Моно, Вайманом и Шанжё[3], или путем последовательной модели (также известной как модель KNF), описанной Кошланд, Неметите и Филмер[4]. Обе модели постулируют, что белковые субъединицы существуют в одной из двух конформаций, напряженной (T-tensed) или расслабленной (R-relaxed), и что расслабленные субъединицы связываются с субстратом легче, чем субъединицы в напряженном состоянии. Эти две модели больше всего различаются своими предположениями о взаимодействии субъединиц и предсуществовании обоих состояний. Для белков, в которых субъединицы существуют в более чем двух конформациях, может использоваться ландшафтная модель аллостерии, описанная Куэнде, Вайнштейном и ЛеВином[5].
Согласованная модель
Согласованная модель аллостерии, также называемая моделью симметрии или моделью MWC, постулирует, что субъединицы фермента связаны таким образом, что конформационные изменения в одной субъединице обязательно передаются всем другим субъединицам. Таким образом, все субъединицы должны существовать в одной и той же конформации. Модель также утверждает, что в отсутствие какого-либо лиганда (субстрата или другого) равновесие благоприятствует одному из конформационных состояний, T или R. Равновесие может быть смещено в состояние R или T посредством связывания одного лиганда (аллостерический эффектор или лиганд) на сайт, отличный от активного сайта (аллостерический сайт).
Последовательная модель
Последовательная модель аллостерической регуляции утверждает, что субъединицы не связаны таким образом, чтобы конформационные изменения в одной вызывали аналогичные изменения в других. Таким образом, все субъединицы фермента не требуют одинаковой конформации. Более того, последовательная модель диктует, что молекулы субстрата связываются посредством протокола индуцированной подгонки. Хотя такая индуцированная подгонка переводит субъединицу из напряженного состояния в расслабленное состояние, она не распространяет конформационные изменения на соседние субъединицы. Вместо этого связывание с субстратом одной субъединицы лишь слегка изменяет структуру других субъединиц, так что их сайты связывания более восприимчивы к субстрату.
Подводя итог:
- субъединицы не обязательно должны существовать в одной и той же конформации
- молекулы субстрата связываются по протоколу индуцированной подгонки
- конформационные изменения не распространяются на все субъединицы
Модель Морфеина
Морфеиновая модель аллостерической регуляции является диссоциативной согласованной моделью[6].
Морфеин — это гомоолигомерная структура, которая может существовать как ансамбль физиологически значимых и функционально различных альтернативных четвертичных ансамблей. Переходы между альтернативными сборками морфеина включают диссоциацию олигомеров, конформационные изменения в диссоциированном состоянии и повторную сборку с образованием другого олигомера. Требуемый этап разборки олигомера отличает модель морфеина для аллостерической регуляции от классических моделей MWC и KNF.
Порфобилиногенсинтаза (PBGS) является прототипом морфеина.
Ансамблевые модели
Ансамблевые модели аллостерической регуляции перечисляют статистический ансамбль аллостерической системы в зависимости от её функции потенциальной энергии, а затем связывают конкретные статистические измерения аллостерии с определёнными энергетическими условиями в энергетической функции (такими как межмолекулярный солевой мост между двумя доменами)[7]. Ансамблевые модели, такие как ансамблевая аллостерическая модель[8] и аллостерическая модель Изинга[9] предполагают, что каждая область системы может принимать два состояния, аналогичные модели MWC. Модель ландшафта аллостерии, введенная Куэндет, Вайнштейном и ЛеВином[5] позволяет доменам иметь любое количество состояний, и вклад определённого молекулярного взаимодействия в данное аллостерическое взаимодействие можно оценить с помощью строгого набора правил. Моделирование молекулярной динамики можно использовать для оценки статистического ансамбля системы, чтобы его можно было проанализировать с помощью ландшафтной модели аллостерии.
Аллостерическая модуляция
Аллостерическая модуляция используется для изменения активности молекул и ферментов в биохимии и фармакологии. Для сравнения, типичное лекарство создается для связывания с активным сайтом фермента, что, таким образом, запрещает связывание субстрата с этим ферментом, вызывая снижение активности фермента. Аллостерическая модуляция происходит, когда эффектор связывается с аллостерическим сайтом (также известным как регуляторный сайт) фермента и изменяет активность фермента. Аллостерические модуляторы разработаны так, чтобы соответствовать аллостерическому сайту, чтобы вызвать конформационное изменение фермента, в частности, изменение формы активного сайта, которое затем вызывает изменение его активности. В отличие от обычных лекарств модуляторы не являются конкурентными ингибиторами. Они могут быть положительными (активирующими), вызывающими повышение активности фермента, или отрицательными (ингибирующими), вызывающими снижение активности фермента. Использование аллостерической модуляции позволяет контролировать эффекты специфической активности ферментов; в результате аллостерические модуляторы очень эффективны в фармакологии[10]. В биологической системе аллостерическую модуляцию трудно отличить от модуляции посредством представления субстрата.
Модель измерения энергии
Примером этой модели является Mycobacterium tuberculosis, бактерия, которая идеально приспособлена к жизни в макрофагах человека. Сайты фермента служат связью между различными субстратами. В частности, между AMФ и Г-6-Ф. Такие сайты также служат механизмом определения эффективности фермента[11].
Положительная модуляция
Положительная аллостерическая модуляция (также известная как аллостерическая активация) происходит, когда связывание одного лиганда увеличивает притяжение между молекулами субстрата и другими сайтами связывания. Примером является связывание молекул кислорода с гемоглобином, где кислород эффективно является и субстратом, и эффектором. Аллостерический или «другой» сайт является активным сайтом прилегающей белковой субъединицы. Связывание кислорода с одной субъединицей вызывает конформационные изменения в этой субъединице, которая взаимодействует с оставшимися активными центрами для повышения их сродства к кислороду. Другой пример аллостерической активации наблюдается в цитозольной IMP-GMP специфической 5'-нуклеотидазе II (cN-II), где сродство к субстрату GMP увеличивается при связывании GTP на границе раздела димеров.
Отрицательная модуляция
Отрицательная аллостерическая модуляция (также известная как аллостерическое ингибирование) происходит, когда связывание одного лиганда снижает сродство к субстрату на других активных сайтах. Например, когда 2,3-БФГ связывается с аллостерическим участком гемоглобина, сродство к кислороду всех субъединиц снижается. Это происходит, когда регулятор отсутствует на сайте привязки.
Прямые ингибиторы тромбина являются прекрасным примером отрицательной аллостерической модуляции. Были обнаружены аллостерические ингибиторы тромбина, которые потенциально могут использоваться в качестве антикоагулянтов.
Другой пример — стрихнин, судорожный яд, который действует как аллостерический ингибитор рецептора глицина. Глицин является основным ингибирующим нейротрансмиттером в спинном мозге и стволе головного мозга млекопитающих. Стрихнин действует в отдельном сайте связывания на рецепторе глицина аллостерическим образом; то есть его связывание снижает сродство рецептора глицина к глицину. Таким образом, стрихнин подавляет действие тормозящего медиатора, что приводит к судорогам.
Другой пример, в котором можно увидеть отрицательную аллостерическую модуляцию, — это связь между АТФ и ферментом фосфофруктокиназой в петле отрицательной обратной связи, которая регулирует гликолиз. Фосфофруктокиназа (обычно называемая ФФК) — это фермент, который катализирует третью стадию гликолиза: фосфорилирование фруктозо-6-фосфата в фруктозо-1,6-бисфосфат. ФФК может аллостерически подавляться высокими уровнями АТФ в клетке. Когда уровни АТФ высоки, АТФ будет связываться с аллосторным участком фосфофруктокиназы, вызывая изменение трехмерной формы фермента. Это изменение вызывает снижение его сродства к субстрату (фруктозо-6-фосфату и АТФ) в активном центре, и фермент становится неактивным. Это приводит к прекращению гликолиза при высоком уровне АТФ, сохраняя глюкозу в организме и поддерживая сбалансированный уровень клеточного АТФ. Таким образом, АТФ служит отрицательным аллостерическим модулятором для ФФК, несмотря на то, что он также является субстратом фермента.
Типы
Гомотропный
Гомотропный аллостерический модулятор является субстратом для своего целевого белка, а также регуляторной молекулой активности белка. Обычно это активатор белка[1]. Например, O2 и CO являются гомеотропными аллостерическими модуляторами гемоглобина. Аналогично, в IMP / GMP-специфической 5'-нуклеотидазе связывание одной молекулы GMP с единственной субъединицей тетрамерного фермента приводит к увеличению сродства к GMP последующих субъединиц, как показано на графиках сигмоидального субстрата в зависимости от скорости.
Гетеротропный
Гетеротропный аллостерический модулятор — это регуляторная молекула, которая не является субстратом фермента. Это может быть активатор или ингибитор фермента. Например, H+, C2 и 2,3-бисфосфоглицерат являются гетеротропными аллостерическими модуляторами гемоглобина[12]. И снова в IMP/GMP-специфической 5'-нуклеотидазе связывание молекулы GTP на границе димера в тетрамерном ферменте приводит к увеличению сродства к субстрату GMP в активном центре, что указывает на гетеротропную аллостерическую активацию K-типа[1].
Как было подробно отмечено выше, некоторые аллостерические белки могут регулироваться как их субстратами, так и другими молекулами. Такие белки способны как к гомотропным, так и к гетеротропным взаимодействиям[1].
Основные активаторы
Некоторые аллостерические активаторы называются «незаменимыми» или «облигатными» активаторами в том смысле, что в их отсутствие активность их целевого фермента очень низкая или незначительная, как в случае с активностью N-ацетилглутамата в отношении карбамоилфосфатсинтетазой I, например[13][14].
Нерегулирующая аллостерия
Нерегулирующий аллостерический сайт — это любой нерегулирующий компонент фермента (или любого белка), который сам по себе не является аминокислотой. Например, многие ферменты требуют связывания натрия для обеспечения правильного функционирования. Однако натрий не обязательно действует как регуляторная субъединица; натрий присутствует всегда, и нет известных биологических процессов добавления/удаления натрия для регулирования активности ферментов. Нерегулирующая аллостерия может содержать любые другие ионы, помимо натрия (кальция, магния, цинка), а также другие химические вещества и, возможно, витамины.
Фармакология
Аллостерическая модуляция рецептора является результатом связывания аллостерических модуляторов в другом сайте (регуляторный сайт), отличном от сайта эндогенного лиганда (активный сайт), и усиливает или ингибирует эффекты эндогенного лиганда. В нормальных условиях он действует, вызывая конформационные изменения в молекуле рецептора, что приводит к изменению аффинности связывания лиганда. Таким образом, аллостерический лиганд модулирует активацию рецептора своим первичным ортостерическим лигандом и, как можно предположить, действует как диммерный переключатель в электрической цепи, регулируя интенсивность ответа.
Например, рецептор ГАМКА имеет два активных сайта, которые связывают нейромедиатор гамма-аминомасляная кислота (ГАМК), но также имеет сайты связывания, регулирующие бензодиазепин и анестетик общего назначения. Каждый из этих регуляторных сайтов может производить положительную аллостерическую модуляцию, усиливая активность ГАМК. Диазепам является положительным аллостерическим модулятором в регуляторном сайте бензодиазепина, а его антидот флумазенил является антагонистом .
Более свежие примеры лекарств, которые аллостерически модулируют свои мишени, включают цинакальцет, имитирующий кальций, и маравирок, применяющийся в лечении ВИЧ.
Аллостерические участки как мишени для лекарств
Аллостерические сайты могут представлять собой новую мишень для лекарств. Существует ряд преимуществ использования аллостерических модуляторов в качестве предпочтительных терапевтических агентов по сравнению с классическими ортостерическими лигандами. Например, сайты аллостерического связывания G-белкового рецептора (GPCR) не столкнулись с таким же эволюционным давлением, как ортостерические сайты для размещения эндогенного лиганда, поэтому они более разнообразны[15]. Следовательно, более высокая селективность GPCR может быть получена путем нацеливания на аллостерические сайты. Это особенно полезно для GPCR, где селективная ортостерическая терапия была затруднена из-за сохранения последовательности ортостерического сайта для всех подтипов рецепторов[16]. Кроме того, эти модуляторы обладают пониженным потенциалом токсических эффектов, поскольку модуляторы с ограниченным взаимодействием будут иметь максимальный уровень своего действия, независимо от введенной дозы. Другой тип фармакологической селективности, который является уникальным для аллостерических модуляторов, основан на кооперативности. Аллостерический модулятор может проявлять нейтральную кооперативность с ортостерическим лигандом на всех подтипах данного рецептора, кроме интересующего подтипа, который называется «абсолютной селективностью подтипа». Если аллостерический модулятор не обладает заметной эффективностью, он может обеспечить другое мощное терапевтическое преимущество по сравнению с ортостерическими лигандами, а именно способность избирательно настраивать повышающие или понижающие тканевые ответы только при наличии эндогенного агониста. Сайты связывания малых молекул, специфичные для олигомеров, являются лекарственными мишенями для имеющих медицинское значение морфеинов[17].
Синтетические аллостерические системы
Существует множество синтетических соединений, содержащих несколько нековалентных сайтов связывания, которые проявляют конформационные изменения при занятии одного сайта. Кооперативность между вкладами единичного связывания в таких супрамолекулярных системах является положительной, если занятие одного сайта связывания увеличивает сродство ΔG во втором сайте, и отрицательной, если сродство не увеличивается. Большинство синтетических аллостерических комплексов основаны на конформационной реорганизации при связывании одного эффекторного лиганда, что затем приводит либо к усилению, либо к ослаблению ассоциации второго лиганда с другим сайтом связывания[18][19][20]. Конформационное связывание между несколькими сайтами связывания в искусственных системах обычно намного больше, чем в белках с их обычно большей гибкостью. Параметр, определяющий эффективность (измеряемый соотношением констант равновесия Krel = KA (E) / KA в присутствии и отсутствии эффектора E), представляет собой конформационную энергию, необходимую для принятия закрытой или напряженной конформации для связывания лиганда А[21].
Во многих поливалентных супрамолекулярных системах[22] может происходить прямое взаимодействие между связанными лигандами, что может привести к большому взаимодействию. Наиболее распространено такое прямое взаимодействие между ионами в рецепторах ионных пар[23][24]. Это сотрудничество часто также называют аллостерией, хотя конформационные изменения здесь не обязательно запускают события связывания.
Интернет-ресурсы
Аллостерическая база данных
Аллостерия — это прямое и эффективное средство регуляции функции биологических макромолекул, возникающее в результате связывания лиганда с аллостерическим сайтом, топографически отличным от ортостерического сайта. Ожидается, что из-за часто высокой селективности рецепторов и более низкой токсичности, связанной с мишенями, аллостерическая регуляция будет играть все большую роль в открытии лекарств и биоинженерии. База данных AlloSteric (ASD, http://mdl.shsmu.edu.cn/ASD)[25] предоставляет центральный ресурс для отображения, поиска и анализа структуры, функций и связанных аннотаций для аллостерических молекул. В настоящее время ASD содержит аллостерические белки более 100 видов и модуляторы трех категорий (активаторы, ингибиторы и регуляторы). Каждый белок снабжен аннотацией с подробным описанием аллостерии, биологического процесса и родственных заболеваний, а каждый модулятор — сродством связывания, физико-химическими свойствами и терапевтической областью. Интеграция информации об аллостерических белках в РАС должна позволить предсказывать аллостерию для неизвестных белков с последующей экспериментальной проверкой. Кроме того, модуляторы, курируемые в ASD, могут использоваться для исследования потенциальных аллостерических мишеней для запрашиваемого соединения и могут помочь химикам реализовать модификации структуры для разработки новых аллостерических лекарств.
Аллостерические аминокислотные остатки и их предсказание
Не все белковые остатки играют одинаково важную роль в аллостерической регуляции. Идентификация остатков, необходимых для аллостерии (так называемых «аллостерических остатков»), была в центре внимания многих исследований, особенно в последнее десятилетие[26][27][28][29][30][31][32][33]. Частично этот растущий интерес является результатом их общей важности в науке о белках, но также и потому, что аллостерические остатки могут использоваться в биомедицинских контекстах. Фармакологически важные белки с сайтами, на которые трудно воздействовать, могут уступить место подходам, в которых один альтернативно нацелен на более легкие для доступа остатки, которые способны аллостерически регулировать первичный интересующий сайт[34]. Эти остатки в широком смысле можно классифицировать как поверхностные и внутренние аллостерические аминокислоты. Аллостерические сайты на поверхности обычно играют регулирующую роль, фундаментально отличную от таковых внутри; поверхностные остатки могут служить рецепторами или эффекторными сайтами при передаче аллостерических сигналов, тогда как те, что находятся внутри, могут действовать в передаче такие сигналы[35].
См. также
- База данных ASD
- Конкурентное ингибирование
- Кооперативное связывание лигандов
- Кинетика ферментов
- Динамика белков
- Теория рецепторов
Примечания
- ↑ 1 2 3 4 Srinivasan B., Forouhar F., Shukla A., Sampangi C., Kulkarni S., Abashidze M., Seetharaman J., Lew S., Mao L., Acton T. B., Xiao R., Everett J. K., Montelione G. T., Tong L., Balaram H. Allosteric regulation and substrate activation in cytosolic nucleotidase II from Legionella pneumophila. (англ.) // The FEBS Journal. — 2014. — March (vol. 281, no. 6). — P. 1613—1628. — doi:10.1111/febs.12727. — PMID 24456211.
- ↑ Proteins move! Protein dynamics and long-range allostery in cell signaling // Protein Structure and Diseases. — 2011. — Vol. 83. — P. 163–221. — ISBN 9780123812629. — doi:10.1016/B978-0-12-381262-9.00005-7.
- ↑ MONOD J, WYMAN J, CHANGEUX JP. ON THE NATURE OF ALLOSTERIC TRANSITIONS: A PLAUSIBLE MODEL. (англ.) // Journal Of Molecular Biology. — 1965. — May (vol. 12). — P. 88—118. — doi:10.1016/s0022-2836(65)80285-6. — PMID 14343300.
- ↑ Koshland Jr. D. E., Némethy G., Filmer D. Comparison of experimental binding data and theoretical models in proteins containing subunits. (англ.) // Biochemistry. — 1966. — January (vol. 5, no. 1). — P. 365—385. — doi:10.1021/bi00865a047. — PMID 5938952.
- ↑ 1 2 Cuendet M. A., Weinstein H., LeVine M. V. The Allostery Landscape: Quantifying Thermodynamic Couplings in Biomolecular Systems. (англ.) // Journal Of Chemical Theory And Computation. — 2016. — 13 December (vol. 12, no. 12). — P. 5758—5767. — doi:10.1021/acs.jctc.6b00841. — PMID 27766843.
- ↑ Jaffe E. K. Morpheeins--a new structural paradigm for allosteric regulation. (англ.) // Trends In Biochemical Sciences. — 2005. — September (vol. 30, no. 9). — P. 490—497. — doi:10.1016/j.tibs.2005.07.003. — PMID 16023348.
- ↑ Motlagh H. N., Wrabl J. O., Li J., Hilser V. J. The ensemble nature of allostery. (англ.) // Nature. — 2014. — 17 April (vol. 508, no. 7496). — P. 331—339. — doi:10.1038/nature13001. — PMID 24740064.
- ↑ Hilser V. J., Wrabl J. O., Motlagh H. N. Structural and energetic basis of allostery. (англ.) // Annual Review Of Biophysics. — 2012. — Vol. 41. — P. 585—609. — doi:10.1146/annurev-biophys-050511-102319. — PMID 22577828.
- ↑ LeVine M. V., Weinstein H. AIM for Allostery: Using the Ising Model to Understand Information Processing and Transmission in Allosteric Biomolecular Systems. (англ.) // Entropy (Basel, Switzerland). — 2015. — May (vol. 17, no. 5). — P. 2895—2918. — doi:10.3390/e17052895. — PMID 26594108.
- ↑ Abdel-Magid A. F. Allosteric modulators: an emerging concept in drug discovery. (англ.) // ACS Medicinal Chemistry Letters. — 2015. — 12 February (vol. 6, no. 2). — P. 104—107. — doi:10.1021/ml5005365. — PMID 25699154.
- ↑ Zhong W., Cui L., Goh B. C., Cai Q., Ho P., Chionh Y. H., Yuan M., Sahili A. E., Fothergill-Gilmore L. A., Walkinshaw M. D., Lescar J., Dedon P. C. Allosteric pyruvate kinase-based "logic gate" synergistically senses energy and sugar levels in Mycobacterium tuberculosis. (англ.) // Nature Communications. — 2017. — 7 December (vol. 8, no. 1). — P. 1986—1986. — doi:10.1038/s41467-017-02086-y. — PMID 29215013.
- ↑ Edelstein S. J. Cooperative interactions of hemoglobin. (англ.) // Annual Review Of Biochemistry. — 1975. — Vol. 44. — P. 209—232. — doi:10.1146/annurev.bi.44.070175.001233. — PMID 237460.
- ↑ Shi D., Allewell N. M., Tuchman M. The N-Acetylglutamate Synthase Family: Structures, Function and Mechanisms. (англ.) // International Journal Of Molecular Sciences. — 2015. — 9 June (vol. 16, no. 6). — P. 13004—13022. — doi:10.3390/ijms160613004. — PMID 26068232.
- ↑ de Cima S., Polo L. M., Díez-Fernández C., Martínez A. I., Cervera J., Fita I., Rubio V. Structure of human carbamoyl phosphate synthetase: deciphering the on/off switch of human ureagenesis. (англ.) // Scientific Reports. — 2015. — 23 November (vol. 5). — P. 16950—16950. — doi:10.1038/srep16950. — PMID 26592762.
- ↑ Christopoulos A., May L. T., Avlani V. A., Sexton P. M. G-protein-coupled receptor allosterism: the promise and the problem(s). (англ.) // Biochemical Society Transactions. — 2004. — November (vol. 32, no. Pt 5). — P. 873—877. — doi:10.1042/BST0320873. — PMID 15494038.
- ↑ May L. T., Leach K., Sexton P. M., Christopoulos A. Allosteric modulation of G protein-coupled receptors. (англ.) // Annual Review Of Pharmacology And Toxicology. — 2007. — Vol. 47. — P. 1—51. — doi:10.1146/annurev.pharmtox.47.120505.105159. — PMID 17009927.
- ↑ Jaffe E. K. MORPHEEINS - A NEW PATHWAY FOR ALLOSTERIC DRUG DISCOVERY. (англ.) // The Open Conference Proceedings Journal. — 2010. — Vol. 1. — P. 1—6. — doi:10.2174/2210289201001010001. — PMID 21643557.
- ↑ Takeuchi M., Ikeda M., Sugasaki A., Shinkai S. Molecular design of artificial molecular and ion recognition systems with allosteric guest responses. (англ.) // Accounts Of Chemical Research. — 2001. — November (vol. 34, no. 11). — P. 865—873. — doi:10.1021/ar0000410. — PMID 11714258.
- ↑ Kremer C., Lützen A. Artificial allosteric receptors. (англ.) // Chemistry (Weinheim An Der Bergstrasse, Germany). — 2013. — 10 May (vol. 19, no. 20). — P. 6162—6196. — doi:10.1002/chem.201203814. — PMID 23463705.
- ↑ Kovbasyuk L., Krämer R. Allosteric supramolecular receptors and catalysts. (англ.) // Chemical Reviews. — 2004. — June (vol. 104, no. 6). — P. 3161—3187. — doi:10.1021/cr030673a. — PMID 15186190.
- ↑ Schneider H. J. Efficiency parameters in artificial allosteric systems. (англ.) // Organic & Biomolecular Chemistry. — 2016. — 14 September (vol. 14, no. 34). — P. 7994—8001. — doi:10.1039/c6ob01303a. — PMID 27431438.
- ↑ Badjić J. D., Nelson A., Cantrill S. J., Turnbull W. B., Stoddart J. F. Multivalency and cooperativity in supramolecular chemistry. (англ.) // Accounts Of Chemical Research. — 2005. — September (vol. 38, no. 9). — P. 723—732. — doi:10.1021/ar040223k. — PMID 16171315.
- ↑ Kim S. K., Sessler J. L. Ion pair receptors. (англ.) // Chemical Society Reviews. — 2010. — October (vol. 39, no. 10). — P. 3784—3809. — doi:10.1039/c002694h. — PMID 20737073.
- ↑ McConnell A. J., Beer P. D. Heteroditopic receptors for ion-pair recognition. (англ.) // Angewandte Chemie (International Ed. In English). — 2012. — 21 May (vol. 51, no. 21). — P. 5052—5061. — doi:10.1002/anie.201107244. — PMID 22419667.
- ↑ Huang Z., Zhu L., Cao Y., Wu G., Liu X., Chen Y., Wang Q., Shi T., Zhao Y., Wang Y., Li W., Li Y., Chen H., Chen G., Zhang J. ASD: a comprehensive database of allosteric proteins and modulators. (англ.) // Nucleic Acids Research. — 2011. — January (vol. 39). — P. D663—669. — doi:10.1093/nar/gkq1022. — PMID 21051350.
- ↑ Panjkovich A., Daura X. Exploiting protein flexibility to predict the location of allosteric sites. (англ.) // BMC Bioinformatics. — 2012. — 25 October (vol. 13). — P. 273—273. — doi:10.1186/1471-2105-13-273. — PMID 23095452.
- ↑ Süel G. M., Lockless S. W., Wall M. A., Ranganathan R. Evolutionarily conserved networks of residues mediate allosteric communication in proteins. (англ.) // Nature Structural Biology. — 2003. — January (vol. 10, no. 1). — P. 59—69. — doi:10.1038/nsb881. — PMID 12483203.
- ↑ Mitternacht S., Berezovsky I. N. Binding leverage as a molecular basis for allosteric regulation. (англ.) // PLoS Computational Biology. — 2011. — September (vol. 7, no. 9). — P. e1002148—1002148. — doi:10.1371/journal.pcbi.1002148. — PMID 21935347.
- ↑ Gasper P. M., Fuglestad B., Komives E. A., Markwick P. R., McCammon J. A. Allosteric networks in thrombin distinguish procoagulant vs. anticoagulant activities. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2012. — 26 December (vol. 109, no. 52). — P. 21216—21222. — doi:10.1073/pnas.1218414109. — PMID 23197839.
- ↑ Ghosh A., Vishveshwara S. Variations in clique and community patterns in protein structures during allosteric communication: investigation of dynamically equilibrated structures of methionyl tRNA synthetase complexes. (англ.) // Biochemistry. — 2008. — 4 November (vol. 47, no. 44). — P. 11398—11407. — doi:10.1021/bi8007559. — PMID 18842003.
- ↑ Sethi A., Eargle J., Black A. A., Luthey-Schulten Z. Dynamical networks in tRNA:protein complexes. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2009. — 21 April (vol. 106, no. 16). — P. 6620—6625. — doi:10.1073/pnas.0810961106. — PMID 19351898.
- ↑ Vanwart A. T., Eargle J., Luthey-Schulten Z., Amaro R. E. Exploring residue component contributions to dynamical network models of allostery. (англ.) // Journal Of Chemical Theory And Computation. — 2012. — 14 August (vol. 8, no. 8). — P. 2949—2961. — doi:10.1021/ct300377a. — PMID 23139645.
- ↑ Rivalta I., Sultan M. M., Lee N. S., Manley G. A., Loria J. P., Batista V. S. Allosteric pathways in imidazole glycerol phosphate synthase. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2012. — 29 May (vol. 109, no. 22). — P. 1428—1436. — doi:10.1073/pnas.1120536109. — PMID 22586084.
- ↑ Negre CFA, Morzan U. N., Hendrickson H. P., Pal R., Lisi G. P., Loria J. P., Rivalta I., Ho J., Batista V. S. Eigenvector centrality for characterization of protein allosteric pathways. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2018. — 26 December (vol. 115, no. 52). — P. 12201—12208. — doi:10.1073/pnas.1810452115. — PMID 30530700.
- ↑ Clarke D., Sethi A., Li S., Kumar S., Chang RWF, Chen J., Gerstein M. Identifying Allosteric Hotspots with Dynamics: Application to Inter- and Intra-species Conservation. (англ.) // Structure (London, England : 1993). — 2016. — 3 May (vol. 24, no. 5). — P. 826—837. — doi:10.1016/j.str.2016.03.008. — PMID 27066750.
| Активность | |
|---|---|
| Регуляция | |
| Классификация | |
| Типы |
|
Обычно почти сразу, изредка в течении часа.