
| Metionina | ||
|---|---|---|
 | ||
 | ||
| Nombre IUPAC | ||
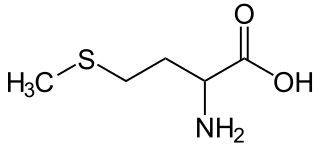
| Ácido 2-amino-4-metiltiobutanoico | ||
| General | ||
| Símbolo químico | Met, M | |
| Fórmula estructural |
 | |
| Fórmula molecular | C5H11NO2S | |
| Identificadores | ||
| Número CAS | 63-68-3[1] | |
| ChEBI | 16811 | |
| ChEMBL | CHEMBL274119 | |
| ChemSpider | 853 | |
| DrugBank | DB00134 | |
| PubChem | 6137 | |
| UNII | 73JWT2K6T3 | |
| KEGG | C01733 D04983, C01733 | |
|
CSCCC(C(=O)O)N
| ||
| Propiedades físicas | ||
| Densidad | 1340 kg/m³; 1,34 g/cm³ | |
| Masa molar | 14 921 g/mol | |
| Punto de fusión | 554 K (281 °C) | |
| Propiedades químicas | ||
| Acidez | 2,16; 9,08 pKa | |
| Solubilidad en agua | soluble | |
| Familia | Aminoácido | |
| Esencial | Sí | |
| Codón | AUG | |
| Punto isoeléctrico (pH) | 5,74 | |
| Valores en el SI y en condiciones estándar (25 ℃ y 1 atm), salvo que se indique lo contrario. | ||
La metionina (abreviada como Met o M) es un aminoácido hidrófobo, cuya fórmula química es: HO2CCH(NH2)CH2CH2SCH3. Al ser hidrófobo este aminoácido esencial está clasificado como no polar.
YouTube Encyclopedic
-
1/5Views:1 438185 3073531 27417 629
-
Metionina: ¿Qué tipo de Aminoácido es?
-
Enlace peptídico / Biología: proteinas
-
TEMA 23. AMINOÁCIDOS Y PÉPTIDOS | 23.3. pH ISOELECTRICO DE CISTEÍNA Y METIONINA
-
Metionina
-
Enlace Peptídico | Formación de Dipéptido y Tripéptido | Aminoácidos | Bioquímica | Mecanismo
Transcription
Historia
El bacteriólogo e inmunólogo estadounidense John Howard Mueller comprobó en 1922 que la adición de una mezcla de los aminoácidos a las colonias de estreptococos (Streptococcus hemolyticus) no era suficiente para su crecimiento.[2] Sin embargo, se logró con la adición de caseína. Por ello Mueller asumió que la caseína todavía debía contener al menos un aminoácido adicional. Durante la investigación posterior de la caseína Mueller por primera vez fue capaz de aislar la metionina.[3][4] La elucidación de la fórmula estructural y de la síntesis la consiguieron en 1926 el británico George Barger y su asistente Frederick Philip Coine,[5] y Barger publicó en 1931 una síntesis mejorada.[6] El nombre de la metionina, una abreviatura de "ácido γ-metiltiol-α-amino-butírico", deriva de un trabajo de S. Odake (1925).[7]
Función
Junto a la cisteína, la metionina es uno de los dos aminoácidos proteinogénicos que contienen azufre. Este deriva de la S-Adenosil metionina (SAM), sirviendo como donante de metilos (la SAM también es usada por las plantas en la síntesis del etileno, en un proceso conocido como ciclo de la metionina o ciclo de Yang).
La metionina es un intermediario en la biosíntesis de la cisteína, la carnitina, la taurina, la lecitina, la fosfatidilcolina y otros fosfolípidos. Fallos en la conversión de metionina pueden desembocar en ateroesclerosis.
Codificación
La metionina es uno de los dos aminoácidos codificados por un único codón del código genético, el AUG (el otro es el triptófano, que está codificado por el codón UGG), que es también el mensaje que indica al ribosoma el inicio de la traducción de una proteína desde el ARNm. Como consecuencia la metionina es el primer aminoácido incorporado, a pesar de que suele ser eliminada en las modificaciones postraduccionales en las diferentes células.
Biosíntesis
Enzimas:
- EC 2.1.1.- Metil Transferasas SAM dependientes
- EC 2.1.1.5 Betaina-homocisteína S-metiltransferasa
- EC 2.1.1.13 Metionina sintasa
- EC 2.3.1.30 Serina acetiltransferasa

Resumen metabólico de la metionina - EC 2.3.1.46 Homoserina O-succiniltransferasa
- EC 2.5.1.6 Metionina adenosiltransferasa
- EC 2.5.1.47 Cisteína sintasa
- EC 2.5.1.48 Cistationina γ-sintasa
- EC 3.3.1.1 S-Adenosilhomocisteína hidrolasa
- EC 4.1.1.57 Metionina descarboxilasa
- EC 4.2.1.22 Cistationina-β-sintasa
- EC 4.4.1.1 Cistationina γ -liasa
- EC 4.4.1.8 Cistationina-β-liasa

Como aminoácido esencial la metionina no es sintetizada en los humanos, por lo tanto hemos de ingerir metionina o proteínas que la contengan. En las plantas y los microorganismos, la metionina es sintetizada por una vía que utiliza tanto ácido aspártico como cisteína. Primero, el ácido aspártico se convierte, vía la β-aspartilo-semialdehído, en homoserina, introduciendo un par de grupos metilenos contiguos. La homoserina pasa a convertirse en 0-succinilhomoserina que tras esto reacciona con la cisteína para producir cistationina que es clave para dar paso a la homocisteína. Posteriormente va la metilación del grupo tiol a partir de fosfatos lo que forma la metionina. Tanto la cistationina-γ-sintetasa y la cistationina-β-sintetasa requieren Piridoxil-5’-fosfato como cofactor, mientras que la metiltransferasa homocisteína requiere de Vitamina B12 como cofactor.
Las enzimas que participan en la biosíntesis de la metionina son:
- Aspartokinasa
- β-aspartato semialdehído deshidrogenasa
- homoserina dehidrogenasa
- homoserina acetiltransferasa
- cistationina-γ-sintetasa
- cistationina-β-liasa
- metionina sintetasa (en mamíferos, este paso se efectúa por la homocisteína metiltransferasa)
Otras vías biomédicas
A pesar de que los mamíferos no pueden sintetizar metionina, aun así todavía se puede utilizar en una gran variedad de vías biomédicas, como en la generación de la homocisteína:
La metionina es convertida a S-adenosilmetionina (SAM) por la metionina adenosiltransferasa, que funciona como donante en muchas reacciones de transferencia de metilos y es convertido en S-adenosilhomocisteína (SAH); la adenocilhomocisteínasa convierte el SAH a homocisteína.
Hay dos destinos de la homocisteína: la regeneración de metionina y la formación de cisteína.
Regeneración de la metionina
La metionina puede ser regenerada a través de la vía de la homocisteína, participando la metionina sintetasa. En esta regeneración desde homocisteína es requerida vitamina B12. Por ello un aumento de homocisteína en análisis clínicos podría ser un signo de deficiencia de esta vitamina.
También puede ser remetilado usando la betaina glicina (NNN-trimetil glicina) a través de la vía de la metionina en la que la enzima beatina-homocisteína metiltransferasa (E.C.2.1.1.5, BHMT). La BHMT representa un 1.5 % de todas las proteínas solubles en el hígado y evidencias recientes sugieren que puede tener una gran influencia en la homeostasis de la metionina y la homocisteína aún mayor que la metionina sintetasa.
Conversión a cisteína
La homocisteína puede ser convertida a cisteína.
- La cistationina-beta-sintetasa (una enzima dependiente del PLP) combina homocisteína y serina para producir cistationina. En vez de degradar cistationina vía cistationina-beta-liasa, característica esta degradación de la biosíntesis, en este caso la cistationina es rota pasando a cisteína y en la α-ketobutirato produciendo cistationina-Y-liasa.
- La alfa-ketoácido deshidrogenada convierte alfa-ketobutirato en propionilo-CoA que es metabolizado a propionil-CoA en un proceso de tres pasos.
Biosíntesis de poliaminas
Las poliaminas espermina y espermidina requieren la transferencia de un sustituyente 3-aminopropil al nitrógeno de una molécula de putrescina o de espermina. Dicho grupo se obtiene por descarboxilación de la S-Adenosil metionina (SAM). Cuando se sintetiza la espermina o la espermidina, el S-metiltiorribósido sufre una serie de transformaciones de tal manera que se recupere la metionina. Dicha ruta se conoce como Ruta de salvamento de la metionina ("Methionine Salvage Pathway").[8]

Enzimas:
- EC 1.13.11.54 Acirreductona dioxigenasa dependiente de hierro (II).
- EC 1.13.11.53 Acirreductona dioxygenasa dependiente de níquel (II)
- EC 2.5.1.22 Espermina sintasa
- EC 2.6.1.5 Tirosina transaminasa (También puede transaminar metionina)[9]
- EC 2.7.1.100 S-Metil-5-tiorribosa quinasa
- EC 3.1.3.77 Acirreductona sintasa
- EC 3.2.2.16 Metiltioadenosina nucleosidasa
- EC 4.1.1.50 S-Adenosilmetionina descarboxilasa
- EC 4.2.1.109 Metiltiorribulosa 1-fosfato deshidratasa
- EC 5.3.1.23 S-metil-5-tiorribosa-1-fosfato isomerasa
Otras biosíntesis
La metionina está implicada en la biosíntesis de etileno,[10] la nicotianamina,[11] las salinosporamidas[12] y varios glucosinolatos[13] tales como la sinigrina, la glucoqueirolina, la glucoerucina, la glucoiberina, la glucoiberverina, la glucorrafanina y el sulforrafano.[14]
Enfermedades metabólicas
La degradación de la metionina está alterada en las siguientes enfermedades metabólicas:
- Acidemia propiónica
- Aciduria malónica y metilmalónica combinada (CMAMMA)
- Aciduria metilmalónica
- Homocistinuria
Fuentes dietéticas
En las semillas de sésamo podemos encontrar niveles bastante altos de metionina, al igual que en nueces brasileñas, pescado, carne y otras semillas de plantas. Existen numerosas frutas y vegetales, así como la mayoría de legumbres, que apenas contienen metionina, sólo en pequeñas cantidades.
La metionina racémica suele añadirse como ingrediente a la comida para mascotas.
| Alimento | g/100g |
|---|---|
| Semillas de sésamo | 1.656 |
| Nueces brasileñas | 1.008 |
| Soja concentrada en proteínas | 0.814 |
| Avena | 0.312 |
| Cacahuetes | 0.309 |
| Garbanzo | 0.253 |
| Maíz | 0.197 |
| Almendra | 0.151 |
| Habas | 0.117 |
| Lentejas | 0.077 |
| Arroz | 0.052 |
Restricción en el consumo de metionina
Cada día son más los estudios que muestran que la restricción en el consumo de metionina puede incrementar el período de vida de algunos animales.[16][17]
En 2005, un estudio mostró que la restricción en el consumo de metionina sin restricción de energía en los roedores, aumenta la duración de su vida.[18]
Véase también
- ARN
- Ciclo de la metionina
- Ribosoma
- Síntesis proteica
Referencias
- ↑ Número CAS
- ↑ S. Hansen: Die Entdeckung der proteinogenen Aminosäuren von 1805 in Paris bis 1935 in Illinois. Archivado el 15 de junio de 2016 en Wayback Machine. Berlin 2015.
- ↑ J. H. Mueller, A new sulphur-containing amino acid isolated from casein., Proc Soc Exp Biol Med, Band 19, S. 161ff (1922).
- ↑ A. M. Pappenheimer, John Howard Mueller, Biographical Memoirs National Academy, 1987, S. 309.
- ↑ G. Barger und F. P. Coine: The amino-acid methionine; constitution and synthesis, Biochem J, Band 22, S. 1417ff (1928).
- ↑ Barger, Weichselbaum, Biochem. J., Band 25, 1931, S. 997.
- ↑ S. Odake, Biochem. Z., Band 161, 1925, 446–455.
- ↑ Jonathan W. Wray, Robert H. Abeles (1995). «The Methionine Salvage Pathway in Klebsiella pneumoniae and Rat Liver IDENTIFICATION AND CHARACTERIZATION OF TWO NOVEL DIOXYGENASES». The Journal of Biological Chemistry, 270: 3147-3153. doi:10.1074/jbc.270.7.3147.
- ↑ Heilbronn J, Wilson J, Berger BJ. (1999). «Tyrosine aminotransferase catalyzes the final step of methionine recycling in Klebsiella pneumoniae.». J Bacteriol. 181 (6): 1739-47.
- ↑ http://www.enzyme-database.org/reaction/misc/ethene.html
- ↑ http://www.enzyme-database.org/reaction/misc/nicotian.html
- ↑ Yolande A. Chan,Angela M. Podevels,Brian M. Kevanya, Michael G. Thomas (2009). «Biosynthesis of polyketide synthase extender units». Natural Product Reports 26: 103. doi:10.1039/b801658p.
- ↑ http://www.plantphysiol.org/cgi/reprint/86/2/319.pdf
- ↑ Paul M. Dewick (2009). Medicinal natural products: a biosynthetic approach. John Wiley and Sons. ISBN 9780470741689.
- ↑ National Nutrient Database for Standard Reference, U.S. Department of Agriculture, archivado desde el original el 3 de marzo de 2015, consultado el 7 de septiembre de 2009..
- ↑ Bárcena C, Quirós PM, Durand S, Mayoral P, Rodríguez F, Caravia XM, Mariño G, Garabaya C, Fernández-García MT, Kroemer G, Freije JMP, López-Otín C (2018). «Methionine Restriction Extends Lifespan in Progeroid Mice and Alters Lipid and Bile Acid Metabolism». PubMed, NCBI. doi:10.1016/j.celrep.2018.07.089.
- ↑ Sun L, Sadighi Akha AA, Miller RA, Harper JM. (2009). «Life-span extension in mice by preweaning food restriction and by methionine restriction in middle age». PubMed, NCBI. PMID 19414512. doi:10.1093/gerona/glp051.
- ↑ Richard A. Miller, Gretchen Buehner, Yayi Chang, James M. Harper, Robert Sigler, Michael Smith‐Wheelock (2005). «Methionine‐deficient diet extends mouse lifespan, slows immune and lens aging, alters glucose, T4, IGF-I and insulin levels, and increases hepatocyte MIF levels and stress resistance». Wiley Online Library. doi:10.1111/j.1474-9726.2005.00152.x.
Enlaces externos
| Control de autoridades |
|
|---|
 Datos: Q180341
Datos: Q180341 Multimedia: Methionine / Q180341
Multimedia: Methionine / Q180341