


La historia genética de los indígenas de América se fundamenta en varios campos, tales como la genética del cromosoma Y, la genética mitocondrial, la genética autosomal y la proteica, los cuales van convergiendo aproximadamente en la misma historia. Los patrones genéticos indican que los indígenas de América experimentaron varios episodios genéticos bien marcados: el primero y más importante se dio con el poblamiento inicial de América proveniente de Siberia, el de los paleoamericanos, el cual sería el factor preponderante en el número de linajes y de marcadores genéticos encontrados en la actual población amerindia. Un poblamiento posterior correspondería al de los pueblos na-dené de Norteamérica y otro al de los esquimo-aleutas en el extremo norte; todos ellos también provenientes de Siberia. Adicionalmente, es posible ―aunque todavía no ha sido comprobado―, el aporte genético europeo en la América precolombina.
YouTube Encyclopedic
-
1/5Views:40 39171 7361 54514 46214 433
-
Los viernes de la evolución: "Historia genómica de los pobladores de América".
-
Reescribiendo la historia humana a través de la genética: el ejemplo de los Agotes
-
El origen del melon: diversidad genética de la especie | 44/87 | UPV
-
El nopal en la frente || El origen genético de los mexicanos
-
La herencia genética de México
Transcription
Hipótesis de las migraciones de Siberia
Antecedentes históricos
Las teorías antropológicas colocaron históricamente a los indígenas americanos en diferentes agrupamientos. En las primeras clasificaciones (François Bernier, siglo XVII) se les clasificó junto con el grupo europeo, en el siglo XVIII solían agruparse independientemente, y en la antropología del siglo XIX comúnmente se relacionaba a indígenas americanos y esquimales con los asiáticos orientales dentro de un grupo mayor denominado raza mongólica.[3]
El primero que determinó específicamente que los indígenas americanos provenían de Asia a través del estrecho de Bering fue el antropólogo checo Aleš Hrdlička (1869-1943), quien sostuvo que los seres humanos americanos se originaron en Mongolia debido al parecido físico y cultural, y que no se realizó este poblamiento de una sola vez, sino que se dio en varias migraciones.[4]
Hipótesis de las tres migraciones
La teoría de las tres migraciones siberianas que poblaron América apareció en 1985, con las primeras investigaciones genéticas[5] y se popularizó a partir de 1986, a partir de los trabajos del lingüista Joseph Greenberg, la paleoantropóloga Christy Turner y el genetista Stephen Zegura, publicando conjuntamente El Poblamiento de América: Una comparación de la evidencia lingüística, dental y genética.[6]
Evidencia lingüística
Joseph Greenberg propuso tres familias principales de lenguas en América: esquimo-aleutianas, na-dené y lenguas amerindias, las cuales equivaldrían a tres procesos distintos del poblamiento de América,[7] aunque sus métodos y conclusiones no son aceptados por la mayoría de lingüistas americanistas (en particular, las críticas se centran en negar el grupo amerindio como grupo filogenéticamente válido desde el punto de vista lingüístico).
Si bien los dos primeros grupos (esquimo-aleutiano, na-dené) son universalmente aceptados y corresponderían a las dos oleadas más recientes, el tercer grupo el amerindio es enormemente diverso y podría corresponder a un proceso migratorio más largo en el que podrían haber participado grupos lingüísticamente diversos, a diferencia del caso de las dos últimas migraciones.
Como antecedentes de la investigación de estos tres grupos se puede citar que las lenguas na-dené fueron establecidas por Edward Sapir en 1915; la relación entre esquimales y aleutas la determinó Rasmus Rusk en 1819, lo cual fue aceptado por lingüistas y antropólogos del siglo XIX y XX; y finalmente los indicios que aparentemente definen las lenguas amerindias fueron enunciados por Alfredo Trombetti en 1905, y este fue respaldado por Sapir en 1918.
La comparación entre los resultados lingüísticos y genéticos es relativa, toda vez que la genética permite obtener conclusiones en base al reloj molecular con miles e incluso millones de años de antigüedad dada la gran variedad de las cadenas nucleicas y proteicas, en cambio la lingüística permite el análisis solo hasta los 5000 o 6000 años de antigüedad, pues en periodos más largos el porcentaje de palabras que muestran el parentesco entre dos lenguas es demasiado bajo para resultar estadísticamente fiable.[8] Si bien es posible que durante el paleolítico se hubiera llevado a cabo más de una migración, sobre la base de la evidencia actual no es posible validar, si bien tampoco descartar la hipótesis amerindia de la única lengua ancestral paleoamericana.[9]
Evidencia antropológica física
La evidencia antropológica dental sugirió que los nativos del norte de China con una primera migración, dieron lugar a la formación de los grupos étnicos paleoamericanos; una segunda migración a los na-dené y una tercera a los esquimo-aleutas.[10]
Análisis genético
Grupo sanguíneo
Estudios de los años 1920 determinaron la predominancia del grupo sanguíneo O en las poblaciones precolombinas, encontrándose también A pero solo en el norte. Jacob Bronowski dedujo en 1973 que en el poblamiento de América se habrían dado al menos dos procesos migratorios: El primero llevando exclusivamente el grupo O, típico de Sudamérica y una segunda ola migratoria trayendo al grupo A solo o acompañado de O, tal como se encuentra en Norteamérica.[11]
Genética proteica
La investigación genética proteica es pionera en el desarrollo de la hipótesis de las tres migraciones. El análisis de la inmunoglobulina G en 1985 reporta una diferente distribución entre los nativos americanos, lo que correspondería según los autores a los tres grupos implicados en el poblamiento de América desde el oriente siberiano y según el siguiente esquema:[5]
| Alotipos mg | Gm1;21 | Gm1,2;21 | Gm1;11,13 |
|---|---|---|---|
| Paleoamericano | Sí | Sí | no |
| Na-dené | Sí | Sí | Sí |
| Esquimo-aleuta | Sí | no | Sí |
En este mismo estudio se encontró el haplotipo Gm3;5,11,13 en bajas frecuencias, el cual es considerado caucásico y de probable origen en el mestizaje.
Genética mitocondrial
Los primeros linajes descubiertos los dio la genética mitocondrial, encontrándose en 1990 cuatro grupos de haplotipos (haplogrupos) en los amerindios[12] y una variante de uno de ellos en los pueblos na-dené.[13] Estos cuatro haplogrupos fueron nombrados en 1992 usando las primeras letras del alfabeto: A, B, C y D, comprobando además el origen asiático de la colonización de América.[14] Al encontrar que en los nativos na-dené dogrib del Canadá se halló casi exclusivamente el grupo A, pronto se dedujo que ello respalda el origen independiente de los pueblos na-dené, pues los amerindios tendrían un origen más antiguo migrando desde Siberia a través del puente de Beringia y con una temprana tribalización.[15]
Pocos años después (1998) se descubrió un quinto linaje, el haplogrupo X, el cual tiene una distribución filogeográfica diferente, ya que mientras los primeros cuatro haplogrupos A, B, C y D se desarrollan en Asia Oriental y se extienden por toda América, X es típico de Eurasia Occidental, encontrándose en Europa en bajas frecuencias y circunscribiéndose en el Nuevo Mundo solo a Norteamérica.[16]
En 2014, el análisis del ADN mitocondrial del esqueleto completo de Naia, datado en 12 900 años AP, encontrado en México, un sistema de cuevas submarinas de Tulum ha probado un vínculo genético entre los paleoamericanos y los modernos nativos americanos ya que encontró que Naia tenía el haplogrupo D1, exclusivo de los actuales amerindios, especialmente de América del Sur.[17]
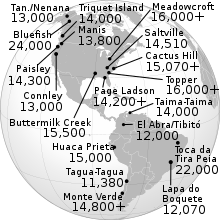
Estos resultados avalan las teorías del poblamiento americano desde Siberia.[18] Las rutas y su antigüedad aproximada se indican en el siguiente mapa:

Genética del cromosoma Y
En los nativos americanos existe un solo linaje patrilineal claramente mayoritario, determinado en 1995, se lo denominó DYS199 (actualmente Q-M3 o Q1a2a1a1) y se presenta en todos los pueblos indígenas americanos, incluidos los esquimales, pero especialmente en Centroamérica y Sudamérica con frecuencias de más del 90%.[19]
Posteriormente se determinaron otros linajes, especialmente los haplogrupos C y R en Norteamérica, por lo que se dedujo que pudo haber dos migraciones primarias procedentes de Siberia,[20] dando lugar a la siguiente distribución:
| Región | Linajes maternos[16] | Linajes paternos[21] |
|---|---|---|
| Norteamérica | A, B, C, D, X | C, Q, R |
| Mesoamérica y Sudamérica | A, B, C, D | Q |
Si bien Q-M3 (Q1a2a1a1, antes Q3) está muy extendido en toda América, su presencia está relacionada específicamente con la primera migración, la de los paleoamericanos. Migraciones posteriores trajeron otros linajes, así pues, en los pueblos na-dené predomina el haplogrupo C-P39 (C2b1a1a, antes C3b1) y en los esquimales Q-NWT01 (Q1a1a).[22]
Genética autosómica
Los últimos estudios genéticos realizados sobre el Polimorfismo de nucleótido concluyen que el poblamiento de América se realizó en tres oleadas migratorias desde Asia, en donde los primeros americanos habrían llegado hace unos 15 000 años y posteriormente llegaron los pueblos na-dené y los esquimoaleutianos.[23]
Principales grupos
El análisis genético lineal subcladístico (en haplogrupos) realizado posteriormente en la población nativa de América, permite identificar grupos y subgrupos cuyo particular acervo genético originario materno (mitocondrial) y paterno (cromosómico Y) puede definirlos.
En la siguiente tabla va en negrita los haplogrupos predominantes y entre paréntesis los que tienen un probable origen en el mestizaje con otros pueblos (orden de norte a sur):
| Grupo | Subrupo | Linajes maternos | Linajes paternos |
|---|---|---|---|
| Esquimo-aleuta | Esquimal | A2a, A2b, D2, D3 | Q-NWT01, Q-M3 (R1). |
| Aleuta | A2a, D2 | Q-M3 (R1). | |
| Na-dené | Na-dené del Norte | A2a (D2). | Q-M3, C-P49 (R1). |
| Na-dené del Sur | A2a (B2, C1). | Q-M3, C-P49 | |
| Paleoamericano | Norteamérica | A2-64, B2, C1, D1, X2 | Q-M3 (C-P49, R1). |
| Mesoamérica y Sudamérica | A2-64, B2, C1, D1, D4h | Q-M3 |
Semejanzas y diferencias
Entre los pueblos indígenas de América, son más las semejanzas que las diferencias. Según la genética autosómica, en el continente americano existe baja diversidad genética y en cambio una gran diferenciación con las poblaciones nativas del resto del mundo.[24] Prueba de esta unidad y uniformidad genética se encuentra en que los linajes fundadores del poblamiento de América A2 (ADNmt) y Q-M3 (ADN-Y), son típicos del continente y con frecuencia predominantes; al igual que el grupo sanguíneo O+.
Las diferencias más sobresalientes serían el resultado de la relación entre las sucesivas olas inmigratorias y la introducción de nuevos linajes. Así pues durante el Paleolítico habrían llegado los primeros linajes paleoamericanos A, B, C, D1 (ADNmt), Q-M3 (ADN-Y) y posiblemente después X (ADNmt); mientras que en el Neolítico vendría C (ADN-Y) posiblemente con los pueblos na-dené, y además D2, D3 (ADNmt) y Q-NWT01 (ADN-Y) con los esquimales.
El último y más detallado estudio sobre el poblamiento de América,[25] analiza 52 pueblos americanos, 17 siberianos y 300 000 variantes de secuencias de ADN, concluyendo en lo siguiente:[26]
- La mayoría de la población nativa americana desciende enteramente de un solo grupo de primeros migrantes que cruzó Beringia, un puente que unió Asia y América durante la era de hielo. De allí se produjo una expansión por todo el continente, seguido de múltiples divergencias y finalmente hubo poco flujo genético entre los grupos nativos americanos, especialmente en América del Sur.
- Se produjeron dos migraciones posteriores que dejaron una huella genética entre los hablantes de lenguas na-dené y esquimoaleutas. Estas migraciones se mezclaron con los primeros pueblos nativos ya establecidos, de tal manera que los pueblos na-dené presentan la mayor parte del genoma de la primera migración, un 90%, mientras que en los esquimoaleutas se conservó el 50 % del mismo.
- También se produjo una migración de regreso de América hacia Siberia, tal como se ve en el análisis genético de los pueblos esquimo-siberianos y en chukchis.
Paleoamericanos

Se denomina paleoamericanos o paleoindios a las poblaciones más antiguas que arribaron a América en el paleolítico (hace más de 10 000 años) y que también son llamados amerindios por muchos genetistas.
En el análisis del cromosoma Y, es común encontrar al haplogrupo Q en frecuencias del 100% en Mesoamérica y Sudamérica, por lo que debe haber sido parte del acervo genético paleoamericano. El mismo fenómeno ocurre con el análisis de grupo sanguíneo, donde el grupo O se encuentra en frecuencias del 100% en muchos grupos étnicos de Iberoamérica.[27]
Paleoantropología genética
El análisis de ADN de los pueblos indígenas actuales, así como el de fósiles humanos, proporciona nuevas luces al estudio del poblamiento de América. Análisis de 42 muestras de las momias chinchorro de hasta 7000 años de antigüedad (de las más antiguas del mundo) dan como resultado A:31,2 B:21,9 C:31,2 y D:3,1% de perfil mitocondrial.[28]
Restos humanos de hace 10 300 años fueron encontrados en Alaska y se informó que pertenecían al haplogrupo mitocondrial D4h3 y al cromosómico Q-M3, los cuales pueden encontrarse actualmente muy extendidos, incluso hasta en Tierra del Fuego.[29] Pero la evidencia genética más antigua la constituye el análisis de coprolitos humanos de hace 12 300 años de las Cuevas de Paisley en Oregón (Estados Unidos), las cuales sustentan el origen paleoamericano preclovis de los amerindios y dieron como resultado A2 y B2 (ADNmt).[30]
Antigüedad
La llegada de los primeros pobladores al continente americano, ha sido ampliamente discutida tanto en sus rutas de procedencia como en su antigüedad, proponiendo fechas tan disímiles como 12 000 o hasta 60 000 años.
Estudios recientes coinciden en que esta colonización se habría producido después del llamado Último Máximo Glacial. Un estudio da una antigüedad de 19 000 años basado en los principales haplogrupos panamericanos A2, B2, C1 y D1;[31] y otro basado en la posibilidad del origen de A2 (ADNmt) en Alaska, le da 17 000 años a la colonización de América desde Siberia.[32] El Proyecto Genográfico afirma que se produjo entre hace 15 000 y 20 000 años.[33]
Según la genética del cromosoma Y la colonización de América rondaría los 14 000 años,[21] según la genética autosomal tiene unos 15 000 años[25] y según el análisis del reloj molecular de todos los grupos mitocondriales americanos, tendría una antigüedad de unos 15 000 años, lo cual coincidiría con las fuentes más fiables del registro arqueológico.[34]
La evidencia arqueológica más confiable afirma que los restos reconocidos como los más antiguos son los de Monte Verde, al sur de Chile, con unos 14 700 años y son más antiguos que los 13 100 de Clovis.[35] Reportes sobre dataciones mucho más antiguas especialmente en Sudamérica, no han sido reconocidos por gran parte de la comunidad científica debido a críticas en los métodos o procedimientos de datación y se mantiene la controversia.[36]
De esta manera todas las evidencias genéticas publicadas en los últimos años avalan la teoría del poblamiento tardío, en contra de un improbable poblamiento temprano que fuera anterior a los 20 000 años. La colonización de todo el continente fue rápida, con una diferencia aproximada de unos 2000 años entre los extremos norteamericano y sudamericano.[37]
Conexión siberiana
Está ampliamente reconocido el poblamiento de América desde Siberia, ya que durante la edad de hielo América y Asia se encontraban unidas a través de un gran puente llamado Beringia.[38]
La evidencia genética refuerza esta tesis, pues relaciona los nativos americanos con los pueblos indígenas de Siberia. Los cuatro haplogrupos panamericanos A, B, C y D (ADNmt) se encuentran bien extendidos en todo el sur de Siberia y marcadamente en buriatos y tuvanos. En los tuvanos por ejemplo se presentan los cuatro grupos sumando un 72% y estableciendo una probable conexión entre los primeros americanos y el sur de Siberia.[39] Sin embargo en el norte de Siberia, donde A y B son escasos, los haplogrupos C y D llegan a sumar 86% en yakutos y evenkis orientales.[40]
Por otro lado, el linaje patrilineal Q (ADN-Y), que es el más común entre todos los pueblos indígenas americanos, tiene en el viejo mundo la mayor frecuencia en Siberia Occidental en los pueblos yeniseos, tales como los ket y selkup.[41]
En conclusión se puede afirmar que si bien hay una conexión entre Siberia y América, no existe en la actualidad una región específica en Eurasia que se relacione absolutamente con el poblamiento de América.[39]
Origen de los ancestros siberianos
Se considera que los primeros emigrantes siberianos que poblaron América tenían a su vez un origen dual, es decir que eran mestizos descendientes de hombres caucásicos y mujeres mongólicas. Pueblos con estas características habitan la Siberia Central en la actualidad.[42]
El análisis genómico de un niño del sur de Siberia de hace 24 000 años confirma esta dualidad, el mestizaje de poblaciones del este de Asia y Eurasia Occidental formó parte del acervo ancestral de los indígenas americanos.[43]
Hipótesis de las rutas colonizadoras paleoamericanas

Hay varias hipótesis sobre las rutas migratorias de los primeros colonizadores, la más aceptada según evidencias geológicas es la hipótesis Clovis del corredor libre de hielo que se habría formado entre Beringia y las Grandes Llanuras de Norteamérica al final de la última glaciación. También es posible una ruta por las costas e islas costeras del Pacífico.
Finalmente se ha sostenido la posibilidad de una migración desde Europa, la hipótesis solutrense, como parte de la herencia ancestral de los amerindios. Sin embargo ninguna de estas tres rutas está totalmente demostrada, si bien tampoco descartada, sobre la base de la evidencia genética.
Hipótesis de la migración única
Según la evidencia mitocondrial, es posible que la colonización paleoamericana fuera el resultado de una sola migración desde Siberia por la ruta costera.[44]
Hipótesis 2 rutas desde Siberia
La idea de dos rutas de colonización paleolítica de América desde Siberia proviene del hecho de que los paleoamericanos pueden dividirse razonablemente en dos subgrupos genéticamente diferenciados, ya sea por el cromosoma-Y o por el ADN mitocondrial o por ambas, generando dos rutas, una muy antigua por la costa Pacífica, y la segunda, hace menos de 12.600 años,[45][46] por el corredor libre de hielo caracterizando dos poblaciones que pueden resumirse del siguiente modo según diversos autores:
| Grupo | Probable ruta[9] | Antigüedad[35] | Linajes maternos[9] | Linajes paternos[21] |
|---|---|---|---|---|
| Amerindios panamericanos | Migración costera del Pacífico | + de 17 500 | A2, B2, C1, C4c, D1, D4h3a[9] | Q |
| Amerindios del este de Norteamérica | Corredor libre de hielo | - de 12 600[46] | A2, B2, C1, D1, X2a, X2g[9] | Q (C3b, R1b). |
Estas teorías se han visto reforzadas por el hallazgo del haplogrupo D4h (también llamado D10) en la isla del Príncipe de Gales (costa del Pacífico de Alaska) de hace 10 300 años.[47] Por otro lado, el hombre de Kennewick presentó los haplogrupos Q-M3 (ADN-Y) y X2a (ADNmt).[48]
Este de Norteamérica: linajes de Occidente
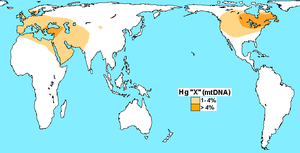
La presencia del haplogrupo X (ADNmt) al este de Norteamérica (ver mapa), un linaje típico de Eurasia Occidental, es coincidente con la presencia de varios clados ADN-Y también típicos de Eurasia Occidental tales como R1b y otros menores (G, J y E).[49] La presencia en América de estos linajes de Occidente ha sido interpretada con las siguientes tres hipótesis:
Origen siberiano
La presencia de X (ADNmt), R1a, R1b y C (ADN-Y) en América, pudo ser el resultado de migraciones de Siberia a través del corredor libre de hielo,[35] ya que todos estos linajes pueden encontrarse, aunque a veces escasamente, en las poblaciones actuales del sur y centro de Siberia.[50][51][52] Esta hipótesis tiene una concordancia étnica y cultural indígena.[20]
Reciente mestizaje europeo
Se considera que la presencia de R1b y otros ADN-Y menores en América sería resultado del mestizaje moderno entre amerindios y europeos.[49] Esta hipótesis sería consistente con la evidencia etnográfica entre el contacto entre los pueblos ojibwa y comerciantes europeos,[53] produciéndose un alto mestizaje similar al de los métis, en donde el linaje materno proviene mayormente de nativas canadienses y el paterno de la colonización anglo-francesa.[54][55] Aunque otros autores apoyan esta hipótesis,[21][56] no se ha realizado ni/o publicado el análisis subcladístico detallado que permita descartar otras hipótesis.
Hipótesis solutrense
Otros autores sostienen la posibilidad de colonización de América por una migración proveniente de Europa, particularmente de Iberia y hace 15 000 años, la cual trajo consigo al haplogrupo X y constituiría una conexión entre la cultura solutrense y la cultura clovis.[57] X es común en toda Europa y particularmente en Irlanda se encontró el linaje X2j, el cual es un clado hermano del linaje americano X2a.[58] Además R1b es predominante en Europa Occidental.[59] Algunos autores han equiparado la industria lítica clovis con la de la cultura solutrense, basados en el estilo de la fabricación de herramientas,[60] pero no se considera que haya evidencia concluyente.[61] También se ha sugerido que los restos del hombre de Kennewick, de unos 7500 años de antigüedad y fisonomía similar a los europeos, podría estar relacionada con una inmigración europea en las primeras etapas del poblamiento de América.[62]
También es posible que esté relacionado con la colonización vikinga en América desde el siglo X y que los nativos skræling de Vinland sean una referencia a los pueblos algonquinos del Canadá;[63][64] o también que pudo existir alguna relación algonquino-celta, toda vez que en los pueblos nórdicos y celtas están presentes estos haplogrupos (X2-ADNmt y R1b-ADN-Y).
Subgrupos amerindios
No existe actualmente una clasificación genética reconocida que permita dividir en subgrupos la población amerindia o paleoamericana. Sin embargo la genética autosómica y la mitocondrial permiten establecer algunas relaciones que coinciden con aspectos lingüísticos y geográficos, pero sin establecer un número bien definido de estos grupos ni fronteras claras entre ellos.
|
Genética autosómicaUn estudio de Cavalli-Sforza y su equipo en 1988,[65] examina por polimorfismo genético a 42 poblaciones alrededor del mundo, concluyendo que hay una correlación entre genética y lingüística de algunas macrofamilias, relacionándo además los tres grandes grupos de nativos americanos con las poblaciones del norte de Asia (Siberia) del siguiente modo (véase el cladograma de la izquierda): |

Un trabajo reciente (2007) de genética autosómica y coordinado entre laboratorios de genética molecular de Estados Unidos, Reino Unido, Suiza, Colombia, Perú, México, Canadá, Brasil, Costa Rica y Chile, relaciona 24 pueblos indígenas americanos con 54 poblaciones del resto del mundo;[24] concluyendo que dentro del continente americano existe baja diversidad genética y en cambio una gran diferenciación con las poblaciones nativas del resto de mundo. Entre los resultados (ver imagen a la derecha) se puede apreciar la mayor divergencia entre los pueblos na-dené (chipewyan) con los amerindios; y a su vez dentro de los amerindios se pueden diferenciar a los pueblos del este de Norteamérica por un lado (los cree y ojibwe, hablantes de lenguas álgicas) con los demás pueblos amerindios por el otro (desde México hasta el extremo sudamericano), formando estos últimos un grupo sólidamente establecido con un 100 % de confiabilidad en su unidad genética. Puede observarse también la estrecha relación genética entre quechuas y aymaras.
Subgrupos mitocondriales y familias lingüísticas

| 1. indígena de Alaska 2. salst 3. atabascano 4. amerindio del norte 5. ojibwa 6. mexica | 7. maya 8. indígena de Centroamérica 9. Andes 10. Amazonia 11. Gran Chaco 12. Patagonia |
Diferentes regiones geográficas presentan una correlación con la forma de distribución de los grandes haplogrupos A, B, C, D y X, los cuales corresponden a su vez con las familias lingüísticas más reconocidas. Las siguientes regiones son las más representativas (de sur a norte):
- Araucanía y Patagonia: Los pueblos del centro y sur de Chile y Argentina se caracterizan por el predominio del haplogrupo D, seguido de C. Se ajustan a ello los hablantes de las llamadas lenguas andinas meridionales tales como mapuches, yaganes, alacalufes[67] y chon.[68]
- Andes: Los pueblos de los Andes presentan gran predominio del haplogrupo B, seguido de D o A, destacando Bolivia seguida del Perú, el norte de Chile, el Ecuador y la región pacífica colombiana. B es mayoritario especialmente en hablantes de lenguas aimaras, uru-chipayas[69] y mosetenas,[70] un poco menos en quechuas[71] y chocó, y no bien definido en barbacoanas.[70] Es posible que en la prehistoria andina no siempre haya predominado B, pues la cultura Chinchorro (Arica) de hace 7000 años presentan un perfil mitocondrial más relacionado con los pueblos del este de Sudamérica.[28]
- Este de Sudamérica: Grupo heterogéneo formado por los pueblos de las regiones de la Amazonía, Gran Chaco y Caribe, incluyendo las Antillas. Los cuatro haplogrupos panamericanos se encuentran dispersos sin que ninguno de ellos sea claramente predominante, aunque sobresale un poco el linaje C, por ejemplo en las familias arahuaca, Caribe, tucana, yanomami, pano-tacana,[72] ticuna, cofán y movima; con una tendencia parcial hacia D en el grupo mataco-guaicurú,[71] hacia B en ye y hacia A en guaraníes.[73]
- Mesoamérica y Centroamérica: En Mesoamérica hay un predominio bien marcado del haplogrupo A, seguido de B, típico de los pueblos mayenses, otomangues y mixe-zoqueanos.[74] Este perfil se extiende por Centroamérica hasta Colombia y es característico en chibchas y paez.[70]
- Aridoamérica: El norte de México y suroeste de los EE. UU. conforman la región de Aridoamérica y en ella prevalece el haplogrupo B. Esta característica es propia de las familias yumano-cochimí, uto-azteca, kiowa-tanoana, caddoana y zuñi, con algunas excepciones como en el caso de los nahuas que poseen un perfil más relacionado con los mesoamericanos.[75]
- Amerindio del norte: El haplogrupo A es mayoritario en todo el este de Norteamérica, norte de Estados Unidos y Canadá. Esto es característico en álgicos, iroqueses, siux, muskogui,[76] salishanos y wakash. Sin embargo lo más particular de esta región es la presencia del haplogrupo X, el cual no tiene una distribución uniforme, pues de lejos su mayor frecuencia y diversidad está en los pueblos algonquinos.[44] La diferencia entre la distribución de X con la del conjunto A-D puede interpretarse como una posible evidencia del origen dual de estos pueblos, en donde X habría arribado posteriormente siendo absorbido por la población inicial.[9]
Pueblos na-dené

Diversos estudios genéticos diferencian a los hablantes de lenguas na-dené de los pueblos esquimo-aleutas y de los amerindios. Pero lo que más caracteriza a los pueblos na-dené es la presencia del haplogrupo C3 (ADN-Y).[77] (ver mapa)
Na-dené del sur y del norte
Una investigación genética proteica (alotipo mg) en el suroeste de EE. UU., encuentra que la diferencia genética más importante en esta región está entre los na-dené del sur (apache, navajo) y los demás pueblos (pima, pápago, hopi, hualapai), reforzando según los autores la tesis de las tres migraciones que poblaron América.[5]
Estudios a partir de catorce locus genéticos encontraron que poblaciones atabascanas (na-dené) están más cercanos a esquimales y chukchis que a los pueblos algonquinos.[78][79] Posteriormente Cavalli-Sforza (1994)[80] hizo un estudio más exhaustivo analizando veintitrés etnias americanas. Como resultado afirmó que hay un distanciamiento de los pueblos na-dené del norte con los del sur, apareciendo los del norte más cercanos a los esquimales, en cambio los na-dené del sur se acercarían a los pueblos amerindios almosanos (álgico-wakash).
Tomando en cuenta el ADN mitocondrial, los na-dené del norte, incluyendo los haida, presentan predominio absoluto del haplogrupo A (ADNmt), mientras que los na-dené del sur presentan otros grupos menores que podrían determinar que se produjo un mestizaje con pueblos amerindios de Norteamérica.[81]
Conexión yenisea

Se ha descubierto afinidad lingüística entre las lenguas na-dené y las lenguas yeniseianas, conformando la familia dené-yenisea.[82]
La evidencia genética del cromosoma-Y nos brinda también una conexión entre estos pueblos. El pueblo siberiano ket, que es prácticamente el único pueblo yeniseo que conserva su lengua original, es también el único que tiene un perfil ADN-Y conformado solo por dos haplogrupos: un mayoritario Q y un minoritario C3,[41] al igual que los apache y otros pueblos na-dené.
En cuanto a la genética mitocondrial, se encontró el linaje materno A2a1 tanto en apaches (na-dené) como en selkupis (yeniseos).[29]
Esquimo-aleutas
Son los hablantes de lenguas esquimo-aleutianas, también llamados escaleutas.[83] La genética mitocondrial ha determinado que son del origen más tardío respecto a los demás grupos nativos americanos, ya que los haplogrupos característicos son relativamente recientes, de 2500 a 5000 años de antigüedad. Los restos más antiguos provienen de Groenlandia, tienen unos 4000 años y han sido analizados genéticamente, lo que permite creer que los primeros esquimo-aleutas habrían llegado de Siberia hace unos 5500 años.[84]
Origen dual
Los linajes característicos escaleutas son D2, D3, A2a, A2b (ADNmt) y Q (ADN-Y). La migración que provino de Siberia trajo consigo los haplogrupos D2 y D3, pero se considera que A2 y sus subclados A2a y A2b son de origen americano.[29] Por lo tanto los pueblos escaleutas tendrían un origen dual: por un lado proveniente de Siberia y por el otro de nativos americanos que colonizaron el Ártico luego que retrocedieran los hielos al terminar la última glaciación. Esta podría ser la razón de la presencia de Q-M3 (ADN-Y) en esquimales.
Conexión chukchi
Igualmente los pueblos chukchi presentan estos mismos haplogrupos D2, D3, A2a y A2b,[32] señal de la estrecha relación genética entre estos pueblos. Ello implicaría además que no solo hubo migraciones siberianas que poblaron América, sino también migraciones americanas que regresaron a Siberia llevando algunos de sus linajes (A2a, A2b y tal vez C1a).[29] La siguiente tabla permite observar esta relación:
| Grupo | Linajes maternos[32] | Linajes paternos |
|---|---|---|
| Inuit | A2a, A2b, D3 | Q-NWT01, Q-M3, R1[22] |
| Aleutas | A2a, D2 | Q-M3 (R1)[85] |
| Esquimo-siberianos | A2a, A2b, D2, D3 | N1c, Q-M3, R1[20] |
| Chukchis | A2a, A2b, C2, C3, D2, D3, G | C3c, N1c, Q-M3, R1 |
Esta conexión genética entre esquimo-aleutas y chukchis podría tener un equivalente lingüístico en una relación entre las lenguas esquimo-aleutianas y las lenguas chucoto-kamchaktas (Swadesh 1962),[86] o incluso relacionarse con la hipótesis de las lenguas uralo-siberianas.
Antigüedad por linajes
Según el ADN mitocondrial
Se ha intentado calcular la antigüedad de cada linaje partiendo de la hipótesis de que a mayor número de mutaciones en el ADN, mayor la edad de cada haplogrupo. Dos estudios recientes basados en el reloj molecular mitocondrial[34][87] dan los siguientes resultados aproximados:

- A: Originado hace unos 30 000 años en Asia Oriental.
- Pre-A2 (o A4): 25 000 años y probable origen chino.
- A2: 13 000[34] o 16 000 años con un probable origen en Alaska[32]
- A2a: 4700, en Norteamérica y Siberia Oriental.[32]
- A2b: 2600, en esquimales y chukchis.
- A2 (64): Con varios subclados presentes en América tales como A2d, A2g, A2h (Colombia), A2i (EE. UU.), A2j, A2k, A2n (Canadá), A2p (Ecuador) y A2r (México y Guatemala).
- A2: 13 000[34] o 16 000 años con un probable origen en Alaska[32]
- Pre-A2 (o A4): 25 000 años y probable origen chino.
- B (16189, 8281) Con 50 000 años, en Asia Oriental.
- B4: 44 000
- B4b: 28 000
- B2: Con 14 600, es autóctono de América.
- B4b: 28 000
- B4: 44 000
- M8: 43 000 años, propio del Extremo Oriente.
- D (16189, 8281) Con 48 000 años, propio del Extremo Oriente.
- D4: 27 000
- D1: 13 500, exclusivo de América.
- D4e: 23 000
- D3 (o D4b): 26 000
- D3a (o D4b1): 20 000.
- D3a2a (o D4b1a2a): En Siberia y esquimales.
- D3a (o D4b1): 20 000.
- D4h: 18 000, en Asia Oriental y en amerindios.[9]
- D4h3b: Típico de los amerindios. Se encontró en la cultura Clovis de hace 12.600 años.[90]
- D4: 27 000
- X (16189, 8281) Con 32 000 años, originado en el Cercano Oriente.
- X2: 21 000
- X2a'j: Además de Norteamérica, también encontrado en Irán, en Siberia (en el Podkamennaya Tunguska),[58] Irlanda y Egipto.
- X2a: 12 800 años y exclusivo de Norteamérica.
- X2a'j: Además de Norteamérica, también encontrado en Irán, en Siberia (en el Podkamennaya Tunguska),[58] Irlanda y Egipto.
- X2: 21 000
Antigüedad según el cromosoma Y
Siendo los linajes más comunes en nativos americanos Q, C y R, la antigüedad aproximada de estos se calcula en lo siguiente:
- C: Originado al sur de Asia hace unos 50 000 años.[77]
- C2: Propio del Extremo Oriente.
- C-P39: Con 14 000 años,[49] típico de los pueblos na-dené (América del Norte).
- C2: Propio del Extremo Oriente.
- P: Originado en el sur de Asia y con unos 34 000 años de antigüedad.[91]
- Q: Con la mayor diversidad en el sur de Asia.
- Q1a1 (F1096): Extendido en el Cercano Oriente y en Extremo Oriente.
- Q1a2 (M346): Extendido en Eurasia, especialmente en Asia Central y Siberia.
- Q1a2a1 (L54) En amerindios y en europeos.
- Q-L54*(xM3) Es un grupo o paragrupo que es el más frecuente entre los amerindios despuéd de Q-M3. Se encontró en la cultura Clovis de hace 12.600 años.[90]
- Q1a2a1a1 (M3): Típico amerindio, con 13 000 años[49] o tal vez 22 000.[92] Fue encontrado en restos humanos de hace 10 300 años en Alaska.[47]
- Q1a2a1 (L54) En amerindios y en europeos.
- R: 27 000[91]
- R1b: Originado en el Cercano Oriente hace 18 500 años, típico de Occidente.
- Q: Con la mayor diversidad en el sur de Asia.
Véase también
- Poblamiento de América
- Poblamiento de América (ruta del Pacífico)
- Poblamiento de América (ruta del Atlántico)
- Paleoamericano
- Prehistoria de América
- América precolombina
- Teoría del poblamiento temprano
- Teoría del poblamiento tardío
- Primeros pobladores andinos
Referencias
- ↑ モンゴロイドの形成 Mapa de las migraciones humanas, en el sitio web del Museo de Kyushu.
- ↑ BURENHULT, Göran (2000). Die ersten menschen. Weltbild Verlag.
- ↑ LESSON, Réné Primeverre (1827). Manuel de mammalogie, ou historie naturelle des mammiferes.
- ↑ HRDLICKA, A. (1942). El origen y la antigüedad de los indios americanos. Washington.
- ↑ a b c WILLIAMS, R. C., et. al. (1985). «GM allotypes in native americans: evidence for three distinct migrations across the Bering land bridge».
- ↑ GREENBERG; TURNER; y ZEGURA (1986). «The settlement of the Americas: a comparison of the linguistic, dental, and genetic evidence».
- ↑ GREENBERG, Joseph (1987). «Languages in the Americas».
- ↑ CAVALLI-SFORZA, Luigi Luca (1992). «Genes, pueblos y lenguas».
- ↑ a b c d e f g PEREGO, Ugo, et. al. (2009). «Distinctive paleo-indian migration routes from Beringia marked by two rare mtDNA haplogroups». En: Current Biology 19, 1, 13 de enero de 2009, págs. 1-8.
- ↑ TURNER, C. G. (1986). «Dentochronological separation estimates for pacific rim populations».
- ↑ BRONOWSKI, Jacob (1975). The ascent of man. Londres: BBC (British Broadcasting Corporation). págs. 92-94. ISBN 0 563 10498 8.
- ↑ SCHURR, T. G., et. al. (1990). «Amerindian mitochondrial DNAs have rare Asian mutations at high frequencies, suggesting they derived from four primary maternal lineages».
- ↑ TORRONI, Antonio, et. al. (1991). «Native american mitochondrial DNA analysis indicates that the amerind and the nadene populations were founded by two independent migrations».
- ↑ WALLACE, Douglas C.; y TORRONI, Antonio (1992). «American indian prehistory as written in the mitochondrial DNA: a review».
- ↑ TORRONI, Antonio, et. al. (1993a). «Afinidades asiáticas y radiación continental de los cuatro grupos fundadores de ADNmt amerindios». Archivado el 24 de septiembre de 2006 en Wayback Machine.
- ↑ a b BROWN, Michael, et. al. (1998). «mtDNA haplogroup X: an ancient link between Europe/Western Asia and North America?».
- ↑
- ↑ «El norte de Asia, cuna de americanos» Archivado el 17 de mayo de 2014 en Wayback Machine.. El Norte de Yucatán (16 de mayo de 2014).
- ↑ UNDERHILL, Peter, et. al. (1995). «A pre-columbian Y chromosome-specific transition and its implications for human evolutionary history».
- ↑ a b c LELL, Jeffrey, et. al. (2001-2002). «The dual origin and siberian affinities of native american».
- ↑ a b c d BORTOLONI, María Catira, et. al. (2003). «Y-chromosome evidence for differing ancient demographic histories in the Americas». Archivado el 28 de abril de 2011 en Wayback Machine.
- ↑ a b c Dulik, Matthew C.; Owings, A. C.; Gaieski, J. B.; Vilar, M. G.; Andre, A.; Lennie, C.; Mackenzie, M. A.; Kritsch, I.; Snowsho, S.; Wright, R.; Martin, J.; Gibson, N.; Andrews, S. D.; Schur, T. G. (2012). «Y-chromosome analysis reveals genetic divergence and new founding native lineages in athapaskan- and eskimoan-speaking populations». En PNAS, 109 (22): págs. 8471-8476.
- ↑ «El estudio genético más detallado sobre el poblamiento de América». Archivado el 17 de julio de 2012 en Wayback Machine. En diario El Comercio (Perú), julio de 2012.
- ↑ a b WANG, Sijia, et. al. 2007, Genetic Variation and Population Structure in Native Americans
- ↑ a b REICH, David, et. al. 2012 Reconstructing Native American population history Nature 2012 DOI: doi:10.1038/nature11258
- ↑ «Native american populations descend from three key migrations, scientists say». En revista Science Daily, julio de 2012.
- ↑ Gran enciclopedia didáctica ilustrada, tomo IX. España: Salvat, 1987.
- ↑ a b Moraga, Mauricio, et. al. (2001). «Análisis de ADN mitocondrial en momias del norte de Chile avala hipótesis de origen amazónico de poblaciones andinas».
- ↑ a b c d e Tamm, Erika, et. al. (2007). «Beringian standstill and spread of native american founders».
- ↑ GILBERT, Thomas, et. al. (2008). «DNA from pre-clovis human coprolites in Oregon, North America». En revista Science, 9, vol. 320, n.º 5877, págs. 786-789, mayo de 2008. DOI: 10.1126/science.1154116
- ↑ ACHILLI, Alessandro, et. al. (2008). «The phylogeny of the four pan-american MtDNA haplogroups: implications for evolutionary and disease studies».
- ↑ a b c d e f Volodko, Natalia, et. al. 2008, Mitochondrial Genome Diversity in Arctic Siberians, with Particular Reference to the Evolutionary History of Beringia and Pleistocenic Peopling of the Americas
- ↑ Atlas de la travesía humana/Marcadores genéticos. En: National Geographic.
- ↑ a b c Soares, Pedro, et. al. (2009). «Correcting for purifying selection: an improved human mitochondrial molecular clock». y su página suplemento. En: The American Journal of Human Genetics, volumen 84, n.º 6, págs. 740-759, 4 de junio de 2009.
- ↑ a b c SCHURR, Theodore; y SHERRY, Stephen (2004). «Mitochondrial DNA and Y chromosome diversity and the peopling of the Americas: evolutionary and demographic evidence».
- ↑ DILLEHAY, Thomas (2000). «Debating the archaeology of the first americans». Archivado el 14 de diciembre de 2014 en Wayback Machine. Capítulo II de The settlement of the Americas.
- ↑ KUMAR, Satish, et. al. (2011). «Large scale mitochondrial sequencing in Mexican Americans suggests a reappraisal of Native American origins». En: BMC Evolutionary Biology, 11: pág. 293, doi:10.1186/1471-2148-11-293
- ↑ HOFFECKER John F.; ELIAS, Scott A. (15 Jun 2007). Human Ecology of Beringia. Columbia University Press. p. 3. ISBN 978-0-231-13060-8.
- ↑ a b Derenko y Malyarchuk 2001, En busca de los ancestros de los nativos americanos (В ПОИСКАХ ПРАРОДИНЫ АМЕРИКАНСКИХ АБОРИГЕНОВ)
- ↑ DERENKO, Miroslava, et. al. (2007). «Phylogeographic analysis of mitochondrial DNA in northern asian populations».
- ↑ a b TAMBETS, Kristiina, et. al. (2004) «The western and eastern roots of the saami—the story of genetic “outliers” told by mitochondrial DNA and Y chromosomes»; The American Journal of Human Genetics 74: 661–682.
- ↑ »Indígenas llegaron a América procedentes de Siberia». En: Gaceta Rusa de Costa Rica, 2010.
- ↑ WADE, Nicholas 2013. 24,000-Year-Old Body Shows Kinship to Europeans and American Indians The New York Times
- ↑ a b Fagundes, N., et. al. (2008). «Mitochondrial Population Genomics Supports a Single Pre-Clovis Origin with a Coastal Route for the Peopling of the Americas» (pdf). American Journal of Human Genetics 82 (3): 583-592. Archivado desde el original el 25 de marzo de 2009. Consultado el 19 de noviembre de 2009.
- ↑ CALLAWAY, Ewen (10 de agosto de 2016). «Plant and animal DNA suggests first Americans took the coastal route». Nature 536 (7615). Consultado el 23 de agosto de 2016.
- ↑ a b PEDERSEN, Mikkel; WILLERSLEV, Eske, et.al. (10 de agosto de 2016). «Postglacial viability and colonization in North America’s ice-free corridor». Nature. Consultado el 23 de agosto de 2016.
- ↑ a b KEMP, Brian, et. al. (2007). «Genetic analysis of early Holocene skeletal remains from Alaska and its implications for the settlement of the Americas».
- ↑ Morten Rasmussen et al. 2015, The ancestry and affiliations of Kennewick Man Nature (2015) doi:10.1038/nature14625
- ↑ a b c d ZEGURA, Stephen, et. al. (2004). «High-resolution SNPs and microsatellite haplotypes point to a single, recent entry of native american Y chromosomes into the Americas».
- ↑ KHAR'KOV, V. N., et. al. (2009), «Comparative characteristics of the gene pool of teleuts inferred from Y-chromosomal marker data». Archivado el 5 de agosto de 2011 en Wayback Machine.
- ↑ DERENKO, M., et. al. (2001), «The presence of mitochondrial haplogroup X in altaians from South Siberia». En American Journal of Human Genetics, 69 (1): págs. 237-241.
- ↑ REIDLA, Maere; KIVISILD, T.; METSPALU, E., et. al. (2003). «Origin and diffusion of mtDNA haplogroup X». En: American Journal of Human Genetics, 73 (5): págs. 1178-1190.
- ↑ RHODES, R. (1982). «Algonquian trade languages». En: COWAN, W. (editor): Proceedings of the 13th Algonquian Conference. Ottawa: Carleton University. págs. 1-10.
- ↑ ««Metis nation of the north west».». Archivado desde el original el 26 de febrero de 2012. Consultado el 31 de julio de 2011.
- ↑ ««Ojibwe history».». Archivado desde el original el 24 de enero de 2013. Consultado el 29 de octubre de 2012.
- ↑ MALHI, Ripan Singh, et. al. (2008). «Distribution of Y chromosomes among native north americans: a study of athapaskan population history».
- ↑ WALLACE, Douglas. «Native American haplogroups: European lineage». Dolan DNA Learning Center.
- ↑ a b REIDLA, Maere; KIVISILD, T.; METSPALU, E., et al. (2003). «Origin and diffusion of mtDNA haplogroup X». En: American Journal of Human Genetics 73 (5): págs. 1178-1190.
- ↑ MC DONALD, J. D. (2005). Mapa de haplogrupos. Archivado el 11 de diciembre de 2013 en Wayback Machine.
- ↑ BRADLEY, B.; y STANFORD, D. (2006). «The solutrean-clovis connection».
- ↑ CARBONELL, Eduard (2006). Homínidos: las primeras ocupaciones de los continentes (pág. 627). Madrid: Ariel.
- ↑ CUSTRED, Glynn. «The forbidden discovery of Kennewick man»
- ↑ «Discovering vikings at L'Anse aux Meadows». Centre for Distance Learning and Innovation, Newfoundland and Labrador, Canadá.
- ↑ Wahlgren, Erik (1990). Los vikingos y América. Barcelona: Destino. 84-233-1915-6.
- ↑ CAVALLI-SFORZA, L. L., et. al. (1988). «Reconstruction of human evolution: Bringing together genetic, archaeological, and linguistic data».
- ↑ «A new genetic map of living humans in interconnected world regions». Archivado el 29 de septiembre de 2011 en Wayback Machine. DNA Tribes.
- ↑ GARCÍA, Federico, et. al. (2004). «Origen y microdiferenciación de la población del archipiélago de Chiloé». (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- ↑ AVENA, Sergio, et. al. (2010). «Mezcla génica y linajes uniparentales en Esquel (provincia de Chubut). Su comparación con otras muestras poblacionales». (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- ↑ SANDOVAL, José, et. al. (2004). «Variantes del ADNmt en isleños del lago Titicaca: máxima frecuencia del haplotipo B1 y evidencia de efecto fundador». (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- ↑ a b c MELTON, Philip, et. al. (2007). «Biological relationship between central and south american chibchan speaking populations: evidence from MtDNA».
- ↑ a b CITLALIN, Xochime (2002). «People of the red brown earth».
- ↑ EASTON, Ruth, et. al. (1996). «mtDNA Variation in the yanomami: evidence for additional new world founding lineages».
- ↑ SALA, Andrea, et. al. (2010). «Genetic analysis of six communities of mbyá-guaraní inhabiting northeastern Argentina by means of nuclear and mitochondrial polymorphic markers».
- ↑ SOLÓRZANO, Eduvigis (2006). «De la Mesoamérica prehispánica a la colonial: la huella del DNA antiguo».
- ↑ MALHI, Ripan, et. al. (2003). «Native american mtDNA prehistory in the american southwest».
- ↑ BOLNICK, Deborah, et. al. (2003). «Unexpected patterns of mitochondrial DNA variation among native americans from the southeastern United States».
- ↑ a b «Dispersión del haplogrupo C». En: National Geographic.
- ↑ SZATHMARY, E. J. E. (1981). «Genetic markers in siberian and northern north american populations». En Yearbook Phys Anthropol, 24: págs. 37-73.
- ↑ SZATHMARY, E. (1985). «Peopling of North America: clues from genetic studies». En: Kirk, R.; Szathmary E. (eds.). «Out of Asia: peopling the Americas and the Pacific». En Journal of Pacific History. Canberra, págs. 79-104.
- ↑ CAVALLI-SFORZA, Luigi Luca (1994). «The history and geography of human genes».
- ↑ mtDNA «Amerind founder haplogroup project - Goals». En: FTDNA Project.
- ↑ VAJDA, Edward (2008). «A siberian link with na-dene languages».
- ↑ DUMOND, Don E. (1965). «On eskaleutian linguistics, archaeology, and prehistory».
- ↑ RASMUSSEN, Morten, et. al. (2009). «Ancient human genome sequence of an extinct palaeo-eskimo».
- ↑ RUBICZ, Rohina Celeste (2007). «Evolutionary consequences of recently founded aleut communities in the Commander and Pribilof Islands».
- ↑ SWADESH, Morris (1962). «Linguistic relations across Behring Strait». En AA, 64: págs. 1262-1291.
- ↑ a b DERENKO, M., et. al. (2010). Origin and Post-Glacial Dispersal of Mitochondrial DNA Haplogroups C and D in Northern Asia
- ↑ PEREGO, Ugo, et. al. (2010). «The initial peopling of the Americas: a growing number of founding mitochondrial genomes from Beringia».
- ↑ GILBERT, Thomas, et. al. (2008). «Paleo-eskimo mtDNA genome reveals matrilineal discontinuity in Greenland».
- ↑ a b Rasmussen M, Anzick SL et al. 2014, The genome of a Late Pleistocene human from a Clovis burial site in western Montana. Nature. 2014 Feb 13;506(7487):225-9. doi: 10.1038/nature13025.
- ↑ a b KARAFET, Tatiana, et. al. (2008). «New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree».
- ↑ BIANCHI, Néstor, et. al. (1997). «Identificación de amerindios por medio del análisis de ADN». (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
Enlaces externos
- América se pobló por primera vez en tres oleadas migratorias desde Asia Noticia de terra.com, julio de 2012
- Poblamiento de América Historias del Mundo. RedEscolar.
- Genética del poblamiento de América. Antropológicas, blog.
- Paleoamerican Origins. Encyclopedia Smithsonian
- Haplogrupos europeos R1b (ADN-Y) y X (ADNmt) en Norteamérica. Forum de Eupedia.com (en inglés).
- Solutrean hypothesis, YouTube
- Haplogrupos del cromosoma Y en los pueblos indígenas de América.
- Indígenas llegaron a América procedentes de Siberia. Revista rusa de Costa Rica.
| Control de autoridades |
|
|---|
 Datos: Q5532907
Datos: Q5532907 Multimedia: Haplogroups / Q5532907
Multimedia: Haplogroups / Q5532907