


Mitochondrial DNA (mtDNA or mDNA)[3] is the DNA located in mitochondria, cellular organelles within eukaryotic cells that convert chemical energy from food into a form that cells can use, such as adenosine triphosphate (ATP). Mitochondrial DNA is only a small portion of the DNA in a eukaryotic cell; most of the DNA can be found in the cell nucleus and, in plants and algae, also in plastids such as chloroplasts.
Human mitochondrial DNA was the first significant part of the human genome to be sequenced.[4] This sequencing revealed that the human mtDNA includes 16,569 base pairs and encodes 13 proteins.
Since animal mtDNA evolves faster than nuclear genetic markers,[5][6][7] it represents a mainstay of phylogenetics and evolutionary biology. It also permits tracing the relationships of populations, and so has become important in anthropology and biogeography.
YouTube Encyclopedic
-
1/5Views:3519 101 72346 379314 4613 855
-
Mitochondrial DNA (mtDNA) Replication - D loop Replication (deep-dive video)
-
DNA Structure and Replication: Crash Course Biology #10
-
"Mitochondrial DNA variation in Human Origins and Disease"
-
Mitochondria Aren't Just the Powerhouse of the Cell
-
Mitochondrial DNA
Transcription
Origin
Nuclear and mitochondrial DNA are thought to have separate evolutionary origins, with the mtDNA derived from the circular genomes of bacteria engulfed by the ancestors of modern eukaryotic cells. This theory is called the endosymbiotic theory. In the cells of extant organisms, the vast majority of the proteins in the mitochondria (numbering approximately 1500 different types in mammals) are coded by nuclear DNA, but the genes for some, if not most, of them are thought to be of bacterial origin, having been transferred to the eukaryotic nucleus during evolution.[8]
The reasons mitochondria have retained some genes are debated. The existence in some species of mitochondrion-derived organelles lacking a genome[9] suggests that complete gene loss is possible, and transferring mitochondrial genes to the nucleus has several advantages.[10] The difficulty of targeting remotely-produced hydrophobic protein products to the mitochondrion is one hypothesis for why some genes are retained in mtDNA;[11] colocalisation for redox regulation is another, citing the desirability of localised control over mitochondrial machinery.[12] Recent analysis of a wide range of mtDNA genomes suggests that both these features may dictate mitochondrial gene retention.[8]
Genome structure and diversity
Across all organisms, there are six main mitochondrial genome types, classified by structure (i.e. circular versus linear), size, presence of introns or plasmid like structures, and whether the genetic material is a singular molecule or collection of homogeneous or heterogeneous molecules.[13]
In many unicellular organisms (e.g., the ciliate Tetrahymena and the green alga Chlamydomonas reinhardtii), and in rare cases also in multicellular organisms (e.g. in some species of Cnidaria), the mtDNA is linear DNA. Most of these linear mtDNAs possess telomerase-independent telomeres (i.e., the ends of the linear DNA) with different modes of replication, which have made them interesting objects of research because many of these unicellular organisms with linear mtDNA are known pathogens.[14]
Animals
Most (bilaterian) animals have a circular mitochondrial genome. Medusozoa and calcarea clades however include species with linear mitochondrial chromosomes.[15] With a few exceptions, animals have 37 genes in their mitochondrial DNA: 13 for proteins, 22 for tRNAs, and 2 for rRNAs.[16]
Mitochondrial genomes for animals average about 16,000 base pairs in length.[16] The anemone Isarachnanthus nocturnus has the largest mitochondrial genome of any animal at 80,923 bp.[17] The smallest known mitochondrial genome in animals belongs to the comb jelly Vallicula multiformis, which consist of 9,961 bp.[18]
In February 2020, a jellyfish-related parasite – Henneguya salminicola – was discovered that lacks a mitochondrial genome but retains structures deemed mitochondrion-related organelles. Moreover, nuclear DNA genes involved in aerobic respiration and in mitochondrial DNA replication and transcription were either absent or present only as pseudogenes. This is the first multicellular organism known to have this absence of aerobic respiration and lives completely free of oxygen dependency.[19][20]
Plants and fungi
There are three different mitochondrial genome types in plants and fungi. The first type is a circular genome that has introns (type 2) and may range from 19 to 1000 kbp in length. The second genome type is a circular genome (about 20–1000 kbp) that also has a plasmid-like structure (1 kb) (type 3). The final genome type found in plants and fungi is a linear genome made up of homogeneous DNA molecules (type 5).[21][22][23]
Great variation in mtDNA gene content and size exists among fungi and plants, although there appears to be a core subset of genes present in all eukaryotes (except for the few that have no mitochondria at all).[8] In Fungi, however, there is no single gene shared among all mitogenomes.[24] Some plant species have enormous mitochondrial genomes, with Silene conica mtDNA containing as many as 11,300,000 base pairs.[25] Surprisingly, even those huge mtDNAs contain the same number and kinds of genes as related plants with much smaller mtDNAs.[26] The genome of the mitochondrion of the cucumber (Cucumis sativus) consists of three circular chromosomes (lengths 1556, 84 and 45 kilobases), which are entirely or largely autonomous with regard to their replication.[27]
Protists
Protists contain the most diverse mitochondrial genomes, with five different types found in this kingdom. Type 2, type 3 and type 5 of the plant and fungal genomes also exist in some protists, as do two unique genome types. One of these unique types is a heterogeneous collection of circular DNA molecules (type 4) while the other is a heterogeneous collection of linear molecules (type 6). Genome types 4 and 6 each range from 1–200 kbp in size.[citation needed]
The smallest mitochondrial genome sequenced to date is the 5,967 bp mtDNA of the parasite Plasmodium falciparum.[28][29]
Endosymbiotic gene transfer, the process by which genes that were coded in the mitochondrial genome are transferred to the cell's main genome, likely explains why more complex organisms such as humans have smaller mitochondrial genomes than simpler organisms such as protists.
| Genome Type[13] | Kingdom | Introns | Size | Shape | Description |
|---|---|---|---|---|---|
| 1 | Animal | No | 11–28 kbp | Circular | Single molecule |
| 2 | Fungi, Plant, Protista | Yes | 19–1000 kbp | Circular | Single molecule |
| 3 | Fungi, Plant, Protista | No | 20–1000 kbp | Circular | Large molecule and small plasmid like structures |
| 4 | Protista | No | 1–200 kbp | Circular | Heterogeneous group of molecules |
| 5 | Fungi, Plant, Protista | No | 1–200 kbp | Linear | Homogeneous group of molecules |
| 6 | Protista | No | 1–200 kbp | Linear | Heterogeneous group of molecules |
Replication
Mitochondrial DNA is replicated by the DNA polymerase gamma complex which is composed of a 140 kDa catalytic DNA polymerase encoded by the POLG gene and two 55 kDa accessory subunits encoded by the POLG2 gene.[30] The replisome machinery is formed by DNA polymerase, TWINKLE and mitochondrial SSB proteins. TWINKLE is a helicase, which unwinds short stretches of dsDNA in the 5' to 3' direction.[31] All these polypeptides are encoded in the nuclear genome.
During embryogenesis, replication of mtDNA is strictly down-regulated from the fertilized oocyte through the preimplantation embryo.[32] The resulting reduction in per-cell copy number of mtDNA plays a role in the mitochondrial bottleneck, exploiting cell-to-cell variability to ameliorate the inheritance of damaging mutations.[33] According to Justin St. John and colleagues, "At the blastocyst stage, the onset of mtDNA replication is specific to the cells of the trophectoderm.[32] In contrast, the cells of the inner cell mass restrict mtDNA replication until they receive the signals to differentiate to specific cell types."[32]

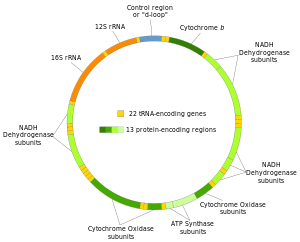
Genes on the human mtDNA and their transcription

The two strands of the human mitochondrial DNA are distinguished as the heavy strand and the light strand. The heavy strand is rich in guanine and encodes 12 subunits of the oxidative phosphorylation system, two ribosomal RNAs (12S and 16S), and 14 transfer RNAs (tRNAs). The light strand encodes one subunit, and 8 tRNAs. So, altogether mtDNA encodes for two rRNAs, 22 tRNAs, and 13 protein subunits, all of which are involved in the oxidative phosphorylation process.[36][37]
| Gene | Type | Product | Positions in the mitogenome |
Strand |
|---|---|---|---|---|
| MT-ATP8 | protein coding | ATP synthase, Fo subunit 8 (complex V) | 08,366–08,572 (overlap with MT-ATP6) | H |
| MT-ATP6 | protein coding | ATP synthase, Fo subunit 6 (complex V) | 08,527–09,207 (overlap with MT-ATP8) | H |
| MT-CO1 | protein coding | Cytochrome c oxidase, subunit 1 (complex IV) | 05,904–07,445 | H |
| MT-CO2 | protein coding | Cytochrome c oxidase, subunit 2 (complex IV) | 07,586–08,269 | H |
| MT-CO3 | protein coding | Cytochrome c oxidase, subunit 3 (complex IV) | 09,207–09,990 | H |
| MT-CYB | protein coding | Cytochrome b (complex III) | 14,747–15,887 | H |
| MT-ND1 | protein coding | NADH dehydrogenase, subunit 1 (complex I) | 03,307–04,262 | H |
| MT-ND2 | protein coding | NADH dehydrogenase, subunit 2 (complex I) | 04,470–05,511 | H |
| MT-ND3 | protein coding | NADH dehydrogenase, subunit 3 (complex I) | 10,059–10,404 | H |
| MT-ND4L | protein coding | NADH dehydrogenase, subunit 4L (complex I) | 10,470–10,766 (overlap with MT-ND4) | H |
| MT-ND4 | protein coding | NADH dehydrogenase, subunit 4 (complex I) | 10,760–12,137 (overlap with MT-ND4L) | H |
| MT-ND5 | protein coding | NADH dehydrogenase, subunit 5 (complex I) | 12,337–14,148 | H |
| MT-ND6 | protein coding | NADH dehydrogenase, subunit 6 (complex I) | 14,149–14,673 | L |
| MT-RNR2 | protein coding | Humanin | — | — |
| MT-TA | transfer RNA | tRNA-Alanine (Ala or A) | 05,587–05,655 | L |
| MT-TR | transfer RNA | tRNA-Arginine (Arg or R) | 10,405–10,469 | H |
| MT-TN | transfer RNA | tRNA-Asparagine (Asn or N) | 05,657–05,729 | L |
| MT-TD | transfer RNA | tRNA-Aspartic acid (Asp or D) | 07,518–07,585 | H |
| MT-TC | transfer RNA | tRNA-Cysteine (Cys or C) | 05,761–05,826 | L |
| MT-TE | transfer RNA | tRNA-Glutamic acid (Glu or E) | 14,674–14,742 | L |
| MT-TQ | transfer RNA | tRNA-Glutamine (Gln or Q) | 04,329–04,400 | L |
| MT-TG | transfer RNA | tRNA-Glycine (Gly or G) | 09,991–10,058 | H |
| MT-TH | transfer RNA | tRNA-Histidine (His or H) | 12,138–12,206 | H |
| MT-TI | transfer RNA | tRNA-Isoleucine (Ile or I) | 04,263–04,331 | H |
| MT-TL1 | transfer RNA | tRNA-Leucine (Leu-UUR or L) | 03,230–03,304 | H |
| MT-TL2 | transfer RNA | tRNA-Leucine (Leu-CUN or L) | 12,266–12,336 | H |
| MT-TK | transfer RNA | tRNA-Lysine (Lys or K) | 08,295–08,364 | H |
| MT-TM | transfer RNA | tRNA-Methionine (Met or M) | 04,402–04,469 | H |
| MT-TF | transfer RNA | tRNA-Phenylalanine (Phe or F) | 00,577–00,647 | H |
| MT-TP | transfer RNA | tRNA-Proline (Pro or P) | 15,956–16,023 | L |
| MT-TS1 | transfer RNA | tRNA-Serine (Ser-UCN or S) | 07,446–07,514 | L |
| MT-TS2 | transfer RNA | tRNA-Serine (Ser-AGY or S) | 12,207–12,265 | H |
| MT-TT | transfer RNA | tRNA-Threonine (Thr or T) | 15,888–15,953 | H |
| MT-TW | transfer RNA | tRNA-Tryptophan (Trp or W) | 05,512–05,579 | H |
| MT-TY | transfer RNA | tRNA-Tyrosine (Tyr or Y) | 05,826–05,891 | L |
| MT-TV | transfer RNA | tRNA-Valine (Val or V) | 01,602–01,670 | H |
| MT-RNR1 | ribosomal RNA | Small subunit : SSU (12S) | 00,648–01,601 | H |
| MT-RNR2 | ribosomal RNA | Large subunit : LSU (16S) | 01,671–03,229 | H |
Between most (but not all) protein-coding regions, tRNAs are present (see the human mitochondrial genome map). During transcription, the tRNAs acquire their characteristic L-shape that gets recognized and cleaved by specific enzymes. With the mitochondrial RNA processing, individual mRNA, rRNA, and tRNA sequences are released from the primary transcript.[39] Folded tRNAs therefore act as secondary structure punctuations.[40]
Regulation of transcription
The promoters for the initiation of the transcription of the heavy and light strands are located in the main non-coding region of the mtDNA called the displacement loop, the D-loop.[36] There is evidence that the transcription of the mitochondrial rRNAs is regulated by the heavy-strand promoter 1 (HSP1), and the transcription of the polycistronic transcripts coding for the protein subunits are regulated by HSP2.[36]
Measurement of the levels of the mtDNA-encoded RNAs in bovine tissues has shown that there are major differences in the expression of the mitochondrial RNAs relative to total tissue RNA.[41] Among the 12 tissues examined the highest level of expression was observed in heart, followed by brain and steroidogenic tissue samples.[41]
As demonstrated by the effect of the trophic hormone ACTH on adrenal cortex cells, the expression of the mitochondrial genes may be strongly regulated by external factors, apparently to enhance the synthesis of mitochondrial proteins necessary for energy production.[41] Interestingly, while the expression of protein-encoding genes was stimulated by ACTH, the levels of the mitochondrial 16S rRNA showed no significant change.[41]
Mitochondrial inheritance
In most multicellular organisms, mtDNA is inherited from the mother (maternally inherited). Mechanisms for this include simple dilution (an egg contains on average 200,000 mtDNA molecules, whereas a healthy human sperm has been reported to contain on average 5 molecules),[42][43] degradation of sperm mtDNA in the male genital tract and in the fertilized egg; and, at least in a few organisms, failure of sperm mtDNA to enter the egg. Whatever the mechanism, this single parent (uniparental inheritance) pattern of mtDNA inheritance is found in most animals, most plants and also in fungi.[44]
In a study published in 2018, human babies were reported to inherit mtDNA from both their fathers and their mothers resulting in mtDNA heteroplasmy,[45] a finding that has been rejected by other scientists.[46][47][48]
Female inheritance
In sexual reproduction, mitochondria are normally inherited exclusively from the mother; the mitochondria in mammalian sperm are usually destroyed by the egg cell after fertilization. Also, mitochondria are only in the sperm tail, which is used for propelling the sperm cells and sometimes the tail is lost during fertilization. In 1999 it was reported that paternal sperm mitochondria (containing mtDNA) are marked with ubiquitin to select them for later destruction inside the embryo.[49] Some in vitro fertilization techniques, particularly injecting a sperm into an oocyte, may interfere with this.
The fact that mitochondrial DNA is mostly maternally inherited enables genealogical researchers to trace maternal lineage far back in time. (Y-chromosomal DNA, paternally inherited, is used in an analogous way to determine the patrilineal history.) This is usually accomplished on human mitochondrial DNA by sequencing the hypervariable control regions (HVR1 or HVR2), and sometimes the complete molecule of the mitochondrial DNA, as a genealogical DNA test.[50] HVR1, for example, consists of about 440 base pairs. These 440 base pairs are compared to the same regions of other individuals (either specific people or subjects in a database) to determine maternal lineage. Most often, the comparison is made with the revised Cambridge Reference Sequence. Vilà et al. have published studies tracing the matrilineal descent of domestic dogs from wolves.[51] The concept of the Mitochondrial Eve is based on the same type of analysis, attempting to discover the origin of humanity by tracking the lineage back in time.
The mitochondrial bottleneck
Entities subject to uniparental inheritance and with little to no recombination may be expected to be subject to Muller's ratchet, the accumulation of deleterious mutations until functionality is lost. Animal populations of mitochondria avoid this through a developmental process known as the mtDNA bottleneck. The bottleneck exploits random processes in the cell to increase the cell-to-cell variability in mutant load as an organism develops: a single egg cell with some proportion of mutant mtDNA thus produces an embryo in which different cells have different mutant loads. Cell-level selection may then act to remove those cells with more mutant mtDNA, leading to a stabilisation or reduction in mutant load between generations. The mechanism underlying the bottleneck is debated,[52][53][54][55] with a recent mathematical and experimental metastudy providing evidence for a combination of the random partitioning of mtDNAs at cell divisions and the random turnover of mtDNA molecules within the cell.[33]
Male inheritance
Male mitochondrial DNA inheritance has been discovered in Plymouth Rock chickens.[56] Evidence supports rare instances of male mitochondrial inheritance in some mammals as well. Specifically, documented occurrences exist for mice,[57][58] where the male-inherited mitochondria were subsequently rejected. It has also been found in sheep,[59] and in cloned cattle.[60] Rare cases of male mitochondrial inheritance have been documented in humans.[61][62][63][45] Although many of these cases involve cloned embryos or subsequent rejection of the paternal mitochondria, others document in vivo inheritance and persistence under lab conditions.
Doubly uniparental inheritance of mtDNA is observed in bivalve mollusks. In those species, females have only one type of mtDNA (F), whereas males have F type mtDNA in their somatic cells, but M type of mtDNA (which can be as much as 30% divergent) in germline cells.[64] Paternally inherited mitochondria have additionally been reported in some insects such as fruit flies,[65][66] honeybees,[67] and periodical cicadas.[68]
Mitochondrial donation
An IVF technique known as mitochondrial donation or mitochondrial replacement therapy (MRT) results in offspring containing mtDNA from a donor female, and nuclear DNA from the mother and father. In the spindle transfer procedure, the nucleus of an egg is inserted into the cytoplasm of an egg from a donor female which has had its nucleus removed, but still contains the donor female's mtDNA. The composite egg is then fertilized with the male's sperm. The procedure is used when a woman with genetically defective mitochondria wishes to procreate and produce offspring with healthy mitochondria.[69] The first known child to be born as a result of mitochondrial donation was a boy born to a Jordanian couple in Mexico on 6 April 2016.[70]
Mutations and disease
Susceptibility
The concept that mtDNA is particularly susceptible to reactive oxygen species generated by the respiratory chain due to its proximity remains controversial.[71] mtDNA does not accumulate any more oxidative base damage than nuclear DNA.[72] It has been reported that at least some types of oxidative DNA damage are repaired more efficiently in mitochondria than they are in the nucleus.[73] mtDNA is packaged with proteins which appear to be as protective as proteins of the nuclear chromatin.[74] Moreover, mitochondria evolved a unique mechanism which maintains mtDNA integrity through degradation of excessively damaged genomes followed by replication of intact/repaired mtDNA. This mechanism is not present in the nucleus and is enabled by multiple copies of mtDNA present in mitochondria.[75] The outcome of mutation in mtDNA may be an alteration in the coding instructions for some proteins,[76] which may have an effect on organism metabolism and/or fitness.
Genetic illness
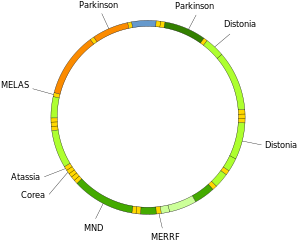
Mutations of mitochondrial DNA can lead to a number of illnesses including exercise intolerance and Kearns–Sayre syndrome (KSS), which causes a person to lose full function of heart, eye, and muscle movements. Some evidence suggests that they might be major contributors to the aging process and age-associated pathologies.[77] Particularly in the context of disease, the proportion of mutant mtDNA molecules in a cell is termed heteroplasmy. The within-cell and between-cell distributions of heteroplasmy dictate the onset and severity of disease[78] and are influenced by complicated stochastic processes within the cell and during development.[33][79]
Mutations in mitochondrial tRNAs can be responsible for severe diseases like the MELAS and MERRF syndromes.[80]
Mutations in nuclear genes that encode proteins that mitochondria use can also contribute to mitochondrial diseases. These diseases do not follow mitochondrial inheritance patterns, but instead follow Mendelian inheritance patterns.[81]
Use in disease diagnosis
Recently a mutation in mtDNA has been used to help diagnose prostate cancer in patients with negative prostate biopsy.[82][83] mtDNA alterations can be detected in the bio-fluids of patients with cancer.[84] mtDNA is characterized by the high rate of polymorphisms and mutations. Some of which are increasingly recognized as an important cause of human pathology such as oxidative phosphorylation (OXPHOS) disorders, maternally inherited diabetes and deafness (MIDD), Type 2 diabetes mellitus, Neurodegenerative disease, heart failure and cancer.
Relationship with ageing
Though the idea is controversial, some evidence suggests a link between aging and mitochondrial genome dysfunction.[85] In essence, mutations in mtDNA upset a careful balance of reactive oxygen species (ROS) production and enzymatic ROS scavenging (by enzymes like superoxide dismutase, catalase, glutathione peroxidase and others). However, some mutations that increase ROS production (e.g., by reducing antioxidant defenses) in worms increase, rather than decrease, their longevity.[71] Also, naked mole rats, rodents about the size of mice, live about eight times longer than mice despite having reduced, compared to mice, antioxidant defenses and increased oxidative damage to biomolecules.[86] Once, there was thought to be a positive feedback loop at work (a 'Vicious Cycle'); as mitochondrial DNA accumulates genetic damage caused by free radicals, the mitochondria lose function and leak free radicals into the cytosol. A decrease in mitochondrial function reduces overall metabolic efficiency.[87] However, this concept was conclusively disproved when it was demonstrated that mice, which were genetically altered to accumulate mtDNA mutations at accelerated rate do age prematurely, but their tissues do not produce more ROS as predicted by the 'Vicious Cycle' hypothesis.[88] Supporting a link between longevity and mitochondrial DNA, some studies have found correlations between biochemical properties of the mitochondrial DNA and the longevity of species.[89] The application of a mitochondrial-specific ROS scavenger, which lead to a significant longevity of the mice studied,[90] suggests that mitochondria may still be well-implicated in ageing. Extensive research is being conducted to further investigate this link and methods to combat ageing. Presently, gene therapy and nutraceutical supplementation are popular areas of ongoing research.[91][92] Bjelakovic et al. analyzed the results of 78 studies between 1977 and 2012, involving a total of 296,707 participants, and concluded that antioxidant supplements do not reduce all-cause mortality nor extend lifespan, while some of them, such as beta carotene, vitamin E, and higher doses of vitamin A, may actually increase mortality.[93] In a recent study, it was showed that dietary restriction can reverse ageing alterations by affecting the accumulation of mtDNA damage in several organs of rats. For example, dietary restriction prevented age-related accumulation of mtDNA damage in the cortex and decreased it in the lung and testis.[94]
Neurodegenerative diseases
Increased mtDNA damage is a feature of several neurodegenerative diseases.
The brains of individuals with Alzheimer's disease have elevated levels of oxidative DNA damage in both nuclear DNA and mtDNA, but the mtDNA has approximately 10-fold higher levels than nuclear DNA.[95] It has been proposed that aged mitochondria is the critical factor in the origin of neurodegeneration in Alzheimer's disease.[96] Analysis of the brains of AD patients suggested an impaired function of the DNA repair pathway, which would cause reduce the overall quality of mtDNA.[97]
In Huntington's disease, mutant huntingtin protein causes mitochondrial dysfunction involving inhibition of mitochondrial electron transport, higher levels of reactive oxygen species and increased oxidative stress.[98] Mutant huntingtin protein promotes oxidative damage to mtDNA, as well as nuclear DNA, that may contribute to Huntington's disease pathology.[99]
The DNA oxidation product 8-oxoguanine (8-oxoG) is a well-established marker of oxidative DNA damage. In persons with amyotrophic lateral sclerosis (ALS), the enzymes that normally repair 8-oxoG DNA damages in the mtDNA of spinal motor neurons are impaired.[100] Thus oxidative damage to mtDNA of motor neurons may be a significant factor in the etiology of ALS.
Correlation of the mtDNA base composition with animal life spans
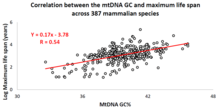
Over the past decade, an Israeli research group led by Professor Vadim Fraifeld has shown that strong and significant correlations exist between the mtDNA base composition and animal species-specific maximum life spans.[101][102][103] As demonstrated in their work, higher mtDNA guanine + cytosine content (GC%) strongly associates with longer maximum life spans across animal species. An additional observation is that the mtDNA GC% correlation with the maximum life spans is independent of the well-known correlation between animal species metabolic rate and maximum life spans. The mtDNA GC% and resting metabolic rate explain the differences in animal species maximum life spans in a multiplicative manner (i.e., species maximum life span = their mtDNA GC% * metabolic rate).[102] To support the scientific community in carrying out comparative analyses between mtDNA features and longevity across animals, a dedicated database was built named MitoAge.[104]
mtDNA mutational spectrum is sensitive to species-specific life-history traits
De novo mutations arise either due to mistakes during DNA replication or due to unrepaired damage caused in turn by endogenous and exogenous mutagens. It has been long believed that mtDNA can be particularly sensitive to damage caused by reactive oxygen species (ROS), however G>T substitutions, the hallmark of the oxidative damage in the nuclear genome, are very rare in mtDNA and do not increase with age. Comparing the mtDNA mutational spectra of hundreds of mammalian species, it has been recently demonstrated that species with extended lifespans have an increased rate of A>G substitutions on single-stranded heavy chain.[105] This discovery led to the hypothesis that A>G is a mitochondria-specific marker of age-associated oxidative damage. This finding provides a mutational (contrary to the selective one) explanation for the observation that long-lived species have GC-rich mtDNA: long-lived species become GC-rich simply because of their biased process of mutagenesis. An association between mtDNA mutational spectrum and species-specific life-history traits in mammals opens a possibility to link these factors together discovering new life-history-specific mutagens in different groups of organisms.
Relationship with non-B (non-canonical) DNA structures
Deletion breakpoints frequently occur within or near regions showing non-canonical (non-B) conformations, namely hairpins, cruciforms and cloverleaf-like elements.[106] Moreover, there is data supporting the involvement of helix-distorting intrinsically curved regions and long G-tetrads in eliciting instability events. In addition, higher breakpoint densities were consistently observed within GC-skewed regions and in the close vicinity of the degenerate sequence motif YMMYMNNMMHM.[107]
Use in forensics
Unlike nuclear DNA, which is inherited from both parents and in which genes are rearranged in the process of recombination, there is usually no change in mtDNA from parent to offspring. Although mtDNA also recombines, it does so with copies of itself within the same mitochondrion. Because of this and because the mutation rate of animal mtDNA is higher than that of nuclear DNA,[108] mtDNA is a powerful tool for tracking ancestry through females (matrilineage) and has been used in this role to track the ancestry of many species back hundreds of generations.
mtDNA testing can be used by forensic scientists in cases where nuclear DNA is severely degraded. Autosomal cells only have two copies of nuclear DNA, but can have hundreds of copies of mtDNA due to the multiple mitochondria present in each cell. This means highly degraded evidence that would not be beneficial for STR analysis could be used in mtDNA analysis. mtDNA may be present in bones, teeth, or hair, which could be the only remains left in the case of severe degradation. In contrast to STR analysis, mtDNA sequencing uses Sanger sequencing. The known sequence and questioned sequence are both compared to the Revised Cambridge Reference Sequence to generate their respective haplotypes. If the known sample sequence and questioned sequence originated from the same matriline, one would expect to see identical sequences and identical differences from the rCRS.[109] Cases arise where there are no known samples to collect and the unknown sequence can be searched in a database such as EMPOP. The Scientific Working Group on DNA Analysis Methods recommends three conclusions for describing the differences between a known mtDNA sequence and a questioned mtDNA sequence: exclusion for two or more differences between the sequences, inconclusive if there is one nucleotide difference, or cannot exclude if there are no nucleotide differences between the two sequences.[110]
The rapid mutation rate (in animals) makes mtDNA useful for assessing genetic relationships of individuals or groups within a species and also for identifying and quantifying the phylogeny (evolutionary relationships; see phylogenetics) among different species. To do this, biologists determine and then compare the mtDNA sequences from different individuals or species. Data from the comparisons is used to construct a network of relationships among the sequences, which provides an estimate of the relationships among the individuals or species from which the mtDNAs were taken. mtDNA can be used to estimate the relationship between both closely related and distantly related species. Due to the high mutation rate of mtDNA in animals, the 3rd positions of the codons change relatively rapidly, and thus provide information about the genetic distances among closely related individuals or species. On the other hand, the substitution rate of mt-proteins is very low, thus amino acid changes accumulate slowly (with corresponding slow changes at 1st and 2nd codon positions) and thus they provide information about the genetic distances of distantly related species. Statistical models that treat substitution rates among codon positions separately, can thus be used to simultaneously estimate phylogenies that contain both closely and distantly related species[80]
Mitochondrial DNA was admitted into evidence for the first time ever in a United States courtroom in 1996 during State of Tennessee v. Paul Ware.[111]
In the 1998 United States court case of Commonwealth of Pennsylvania v. Patricia Lynne Rorrer,[112] mitochondrial DNA was admitted into evidence in the State of Pennsylvania for the first time.[113][114] The case was featured in episode 55 of season 5 of the true crime drama series Forensic Files (season 5).[115]
Mitochondrial DNA was first admitted into evidence in California, United States, in the successful prosecution of David Westerfield for the 2002 kidnapping and murder of 7-year-old Danielle van Dam in San Diego: it was used for both human and dog identification.[116] This was the first trial in the U.S. to admit canine DNA.[117]
The remains of King Richard III, who died in 1485, were identified by comparing his mtDNA with that of two matrilineal descendants of his sister who were alive in 2013, 527 years after he died.[118]
Use in evolutionary biology and systematic biology
mtDNA is conserved across eukaryotic organism given the critical role of mitochondria in cellular respiration. However, due to less efficient DNA repair (compared to nuclear DNA) it has a relatively high mutation rate (but slow compared to other DNA regions such as microsatellites) which makes it useful for studying the evolutionary relationships—phylogeny—of organisms. Biologists can determine and then compare mtDNA sequences among different species and use the comparisons to build an evolutionary tree for the species examined.
For instance, while most nuclear genes are nearly identical between humans and chimpanzees, their mitochondrial genomes are 9.8% different. Human and gorilla mitochondrial genomes are 11.8% different, suggesting that humans may be more closely related to chimpanzees than gorillas.[119]
mtDNA in nuclear DNA
Whole genome sequences of more than 66,000 people revealed that most of them had some mitochondrial DNA inserted into their nuclear genomes. More than 90% of these nuclear-mitochondrial segments (NUMTs) were inserted after humans diverged from apes. Results indicate such transfers currently occur as frequent as once in every ~4,000 human births.[120]
It appears that organellar DNA is much more often transferred to nuclear DNA than previously thought. This observation also supports the idea of the endosymbiont theory that eukaryotes have evolved from endosymbionts which turned into organelles while transferring most of their DNA to the nucleus so that the organellar genome shrunk in the process.[121]
History
Mitochondrial DNA was discovered in the 1960s by Margit M. K. Nass and Sylvan Nass by electron microscopy as DNase-sensitive threads inside mitochondria,[122] and by Ellen Haslbrunner, Hans Tuppy and Gottfried Schatz by biochemical assays on highly purified mitochondrial fractions.[123]
Mitochondrial sequence databases
Several specialized databases have been founded to collect mitochondrial genome sequences and other information. Although most of them focus on sequence data, some of them include phylogenetic or functional information.
- AmtDB: a database of ancient human mitochondrial genomes.[124]
- InterMitoBase: an annotated database and analysis platform of protein-protein interactions for human mitochondria.[125] (apparently last updated in 2010, but still available)
- MitoBreak: the mitochondrial DNA breakpoints database.[126]
- MitoFish and MitoAnnotator: a mitochondrial genome database of fish.[127] See also Cawthorn et al.[128]
- Mitome: a database for comparative mitochondrial genomics in metazoan animals[129] (no longer available)
- MitoRes: a resource of nuclear-encoded mitochondrial genes and their products in metazoa[130] (apparently no longer being updated)
- MitoSatPlant: Mitochondrial microsatellites database of viridiplantae.[131]
- MitoZoa 2.0: a database for comparative and evolutionary analyses of mitochondrial genomes in Metazoa.[132] (no longer available)
MtDNA-phenotype association databases
Genome-wide association studies can reveal associations of mtDNA genes and their mutations with phenotypes including lifespan and disease risks. In 2021, the largest, UK Biobank-based, genome-wide association study of mitochondrial DNA unveiled 260 new associations with phenotypes including lifespan and disease risks for e.g. type 2 diabetes.[133][134]
Mitochondrial mutation databases
Several specialized databases exist that report polymorphisms and mutations in the human mitochondrial DNA, together with the assessment of their pathogenicity.
- MitImpact: A collection of pre-computed pathogenicity predictions for all nucleotide changes that cause non-synonymous substitutions in human mitochondrial protein coding genes MitImpact 3D - IRCCS-CSS Bioinformatics lab.
- MITOMAP: A compendium of polymorphisms and mutations in human mitochondrial DNA WebHome < MITOMAP < Foswiki.
See also
References
- ^ Siekevitz P (1957). "Powerhouse of the cell". Scientific American. 197 (1): 131–40. Bibcode:1957SciAm.197a.131S. doi:10.1038/scientificamerican0757-131.
- ^ Iborra FJ, Kimura H, Cook PR (May 2004). "The functional organization of mitochondrial genomes in human cells". BMC Biology. 2: 9. doi:10.1186/1741-7007-2-9. PMC 425603. PMID 15157274.
- ^ Sykes B (10 September 2003). "Mitochondrial DNA and human history". The Human Genome. Wellcome Trust. Archived from the original on 7 September 2015. Retrieved 5 February 2012.
- ^ Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J, et al. (April 1981). "Sequence and organization of the human mitochondrial genome". Nature. 290 (5806): 457–465. Bibcode:1981Natur.290..457A. doi:10.1038/290457a0. PMID 7219534. S2CID 4355527.
- ^ Boursot P, Bonhomme F (1 January 1986). "[Not Available]". Génétique, Sélection, Évolution. 18 (1): 73–98. doi:10.1186/1297-9686-18-1-73. PMC 2713894. PMID 22879234.
- ^ Delsuc F, Stanhope MJ, Douzery EJ (August 2003). "Molecular systematics of armadillos (Xenarthra, Dasypodidae): contribution of maximum likelihood and Bayesian analyses of mitochondrial and nuclear genes". Molecular Phylogenetics and Evolution. 28 (2): 261–275. doi:10.1016/s1055-7903(03)00111-8. PMID 12878463. Archived from the original on 4 November 2018. Retrieved 4 November 2018.
- ^ Hassanin A, An J, Ropiquet A, Nguyen TT, Couloux A (March 2013). "Combining multiple autosomal introns for studying shallow phylogeny and taxonomy of Laurasiatherian mammals: Application to the tribe Bovini (Cetartiodactyla, Bovidae)". Molecular Phylogenetics and Evolution. 66 (3): 766–775. doi:10.1016/j.ympev.2012.11.003. PMID 23159894.
- ^ a b c Johnston IG, Williams BP (February 2016). "Evolutionary Inference across Eukaryotes Identifies Specific Pressures Favoring Mitochondrial Gene Retention" (PDF). Cell Systems. 2 (2): 101–111. doi:10.1016/j.cels.2016.01.013. PMID 27135164.
- ^ van der Giezen M, Tovar J, Clark CG (2005). "Mitochondrion-Derived Organelles in Protists and Fungi". A Survey of Cell Biology. International Review of Cytology. Vol. 244. pp. 175–225. doi:10.1016/S0074-7696(05)44005-X. ISBN 978-0-12-364648-4. PMID 16157181.
- ^ Adams KL, Palmer JD (December 2003). "Evolution of mitochondrial gene content: gene loss and transfer to the nucleus". Molecular Phylogenetics and Evolution. 29 (3): 380–395. doi:10.1016/S1055-7903(03)00194-5. PMID 14615181.
- ^ Björkholm P, Harish A, Hagström E, Ernst AM, Andersson SG (August 2015). "Mitochondrial genomes are retained by selective constraints on protein targeting". Proceedings of the National Academy of Sciences of the United States of America. 112 (33): 10154–10161. Bibcode:2015PNAS..11210154B. doi:10.1073/pnas.1421372112. PMC 4547212. PMID 26195779.
- ^ Allen JF (August 2015). "Why chloroplasts and mitochondria retain their own genomes and genetic systems: Colocation for redox regulation of gene expression". Proceedings of the National Academy of Sciences of the United States of America. 112 (33): 10231–10238. Bibcode:2015PNAS..11210231A. doi:10.1073/pnas.1500012112. PMC 4547249. PMID 26286985.
- ^ a b Kolesnikov AA, Gerasimov ES (December 2012). "Diversity of mitochondrial genome organization". Biochemistry. Biokhimiia. 77 (13): 1424–1435. doi:10.1134/S0006297912130020. PMID 23379519. S2CID 14441187.
- ^ Nosek J, Tomáska L, Fukuhara H, Suyama Y, Kovác L (May 1998). "Linear mitochondrial genomes: 30 years down the line". Trends in Genetics. 14 (5): 184–188. doi:10.1016/S0168-9525(98)01443-7. PMID 9613202.
- ^ Lavrov DV, Pett W (September 2016). "Animal Mitochondrial DNA as We Do Not Know It: mt-Genome Organization and Evolution in Nonbilaterian Lineages". Genome Biology and Evolution. 8 (9): 2896–2913. doi:10.1093/gbe/evw195. PMC 5633667. PMID 27557826.
- ^ a b Boore, J. L. (1 April 1999). "Animal mitochondrial genomes". Nucleic Acids Research. 27 (8): 1767–1780. doi:10.1093/nar/27.8.1767. PMC 148383. PMID 10101183.
- ^ Stampar SN, Broe MB, Macrander J, Reitzel AM, Brugler MR, Daly M (April 2019). "Linear Mitochondrial Genome in Anthozoa (Cnidaria): A Case Study in Ceriantharia". Scientific Reports. 9 (1): 6094. Bibcode:2019NatSR...9.6094S. doi:10.1038/s41598-019-42621-z. PMC 6465557. PMID 30988357.
- ^ Polymorphism within the mitochondrial genome of the ctenophore, Pleurobrachia bachei and its ongoing rapid evolution - bioRxiv
- ^ Yahalomi D, Atkinson SD, Neuhof M, Chang ES, Philippe H, Cartwright P, et al. (March 2020). "A cnidarian parasite of salmon (Myxozoa: Henneguya) lacks a mitochondrial genome". Proceedings of the National Academy of Sciences of the United States of America. 117 (10): 5358–5363. Bibcode:2020PNAS..117.5358Y. doi:10.1073/pnas.1909907117. PMC 7071853. PMID 32094163.
- ^ Starr M (25 February 2020). "Scientists Find The First-Ever Animal That Doesn't Need Oxygen to Survive". ScienceAlert. Archived from the original on 25 February 2020. Retrieved 25 February 2020.
- ^ de Queiroz, Casley Borges; Santana, Mateus Ferreira; Pereira Vidigal, Pedro M.; de Queiroz, Marisa Vieira (15 March 2018). "Comparative analysis of the mitochondrial genome of the fungus Colletotrichum lindemuthianum, the causal agent of anthracnose in common beans". Applied Microbiology & Biotechnology. 102 (6): 2763–2778. doi:10.1007/s00253-018-8812-0. PMID 29453633. S2CID 3341230.
- ^ Masutani, Bansho; Arimura, Shin-ichi; Morishita, Shinichi (12 January 2021). "Investigating the mitochondrial genomic landscape of Arabidopsis thaliana by long-read sequencing". PLOS Computational Biology. 17 (1): e1008597. Bibcode:2021PLSCB..17E8597M. doi:10.1371/journal.pcbi.1008597. PMC 7833223. PMID 33434206.
- ^ Kozik, Alexander; Rowan, Beth A.; Lavelle, Dean; Berke, Lidija; Schranz, M. Eric; Michelmore, Richard W.; Christensen, Alan C. (30 August 2019). "The alternative reality of plant mitochondrial DNA: One ring does not rule them all". PLOS Genetics. 15 (8): e1008373. doi:10.1371/journal.pgen.1008373. PMC 6742443. PMID 31469821.
- ^ Fonseca PL, De-Paula RB, Araújo DS, Tomé LM, Mendes-Pereira T, Rodrigues WF, et al. (2021). "Global Characterization of Fungal Mitogenomes: New Insights on Genomic Diversity and Dynamism of Coding Genes and Accessory Elements". Frontiers in Microbiology. 12: 787283. doi:10.3389/fmicb.2021.787283. PMC 8672057. PMID 34925295.
- ^ Sloan DB, Alverson AJ, Chuckalovcak JP, Wu M, McCauley DE, Palmer JD, Taylor DR (January 2012). "Rapid evolution of enormous, multichromosomal genomes in flowering plant mitochondria with exceptionally high mutation rates". PLOS Biology. 10 (1): e1001241. doi:10.1371/journal.pbio.1001241. PMC 3260318. PMID 22272183.
- ^ Ward BL, Anderson RS, Bendich AJ (September 1981). "The mitochondrial genome is large and variable in a family of plants (cucurbitaceae)". Cell. 25 (3): 793–803. doi:10.1016/0092-8674(81)90187-2. PMID 6269758. S2CID 45090891.
- ^ Alverson AJ, Rice DW, Dickinson S, Barry K, Palmer JD (July 2011). "Origins and recombination of the bacterial-sized multichromosomal mitochondrial genome of cucumber". The Plant Cell. 23 (7): 2499–2513. doi:10.1105/tpc.111.087189. JSTOR 41433488. PMC 3226218. PMID 21742987.
- ^ "Mitochondrial DNA (mtDNA)" (PDF). Integrated DNA Technologies. Archived from the original (PDF) on 29 July 2016. Retrieved 25 February 2016.
- ^ Tyagi S, Pande V, Das A (February 2014). "Whole mitochondrial genome sequence of an Indian Plasmodium falciparum field isolate". The Korean Journal of Parasitology. 52 (1): 99–103. doi:10.3347/kjp.2014.52.1.99. PMC 3949004. PMID 24623891.
- ^ Yakubovskaya E, Chen Z, Carrodeguas JA, Kisker C, Bogenhagen DF (January 2006). "Functional human mitochondrial DNA polymerase gamma forms a heterotrimer". The Journal of Biological Chemistry. 281 (1): 374–382. doi:10.1074/jbc.M509730200. PMID 16263719.
- ^ Jemt E, Farge G, Bäckström S, Holmlund T, Gustafsson CM, Falkenberg M (November 2011). "The mitochondrial DNA helicase TWINKLE can assemble on a closed circular template and support initiation of DNA synthesis". Nucleic Acids Research. 39 (21): 9238–9249. doi:10.1093/nar/gkr653. PMC 3241658. PMID 21840902.
- ^ a b c St John JC, Facucho-Oliveira J, Jiang Y, Kelly R, Salah R (2010). "Mitochondrial DNA transmission, replication and inheritance: a journey from the gamete through the embryo and into offspring and embryonic stem cells". Human Reproduction Update. 16 (5): 488–509. doi:10.1093/humupd/dmq002. PMID 20231166.
- ^ a b c Johnston IG, Burgstaller JP, Havlicek V, Kolbe T, Rülicke T, Brem G, et al. (June 2015). "Stochastic modelling, Bayesian inference, and new in vivo measurements elucidate the debated mtDNA bottleneck mechanism". eLife. 4: e07464. arXiv:1512.02988. doi:10.7554/eLife.07464. PMC 4486817. PMID 26035426.
- ^ Shuster RC, Rubenstein AJ, Wallace DC (September 1988). "Mitochondrial DNA in anucleate human blood cells". Biochemical and Biophysical Research Communications. 155 (3): 1360–1365. doi:10.1016/s0006-291x(88)81291-9. PMID 3178814.
- ^ Zhang D, Keilty D, Zhang ZF, Chian RC (March 2017). "Mitochondria in oocyte aging: current understanding". Facts, Views & Vision in ObGyn. 9 (1): 29–38. PMC 5506767. PMID 28721182.
- ^ a b c Barshad G, Marom S, Cohen T, Mishmar D (September 2018). "Mitochondrial DNA Transcription and Its Regulation: An Evolutionary Perspective". Trends in Genetics. 34 (9): 682–692. doi:10.1016/j.tig.2018.05.009. PMID 29945721. S2CID 49430452.
- ^ Barchiesi A, Vascotto C (May 2019). "Transcription, Processing, and Decay of Mitochondrial RNA in Health and Disease". International Journal of Molecular Sciences. 20 (9): 2221. doi:10.3390/ijms20092221. PMC 6540609. PMID 31064115.
- ^ Homo sapiens mitochondrion, complete genome. "Revised Cambridge Reference Sequence (rCRS): accession NC_012920" Archived 6 April 2020 at the Wayback Machine, National Center for Biotechnology Information. Retrieved on 20 February 2017.
- ^ Falkenberg M, Larsson NG, Gustafsson CM (19 June 2007). "DNA replication and transcription in mammalian mitochondria". Annual Review of Biochemistry. 76 (1): 679–699. doi:10.1146/annurev.biochem.76.060305.152028. PMID 17408359.
- ^ Ojala D, Montoya J, Attardi G (April 1981). "tRNA punctuation model of RNA processing in human mitochondria". Nature. 290 (5806): 470–474. Bibcode:1981Natur.290..470O. doi:10.1038/290470a0. PMID 7219536. S2CID 4323371.
- ^ a b c d Raikhinstein M, Hanukoglu I (November 1993). "Mitochondrial-genome-encoded RNAs: differential regulation by corticotropin in bovine adrenocortical cells". Proceedings of the National Academy of Sciences of the United States of America. 90 (22): 10509–10513. Bibcode:1993PNAS...9010509R. doi:10.1073/pnas.90.22.10509. PMC 47806. PMID 7504267.
- ^ Wolff JN, Gemmell NJ (November 2008). "Lost in the zygote: the dilution of paternal mtDNA upon fertilization". Heredity. 101 (5): 429–434. doi:10.1038/hdy.2008.74. PMID 18685570. S2CID 5065133.
- ^ Gabriel MS, Chan SW, Alhathal N, Chen JZ, Zini A (August 2012). "Influence of microsurgical varicocelectomy on human sperm mitochondrial DNA copy number: a pilot study". Journal of Assisted Reproduction and Genetics. 29 (8): 759–764. doi:10.1007/s10815-012-9785-z. PMC 3430774. PMID 22562241.
- ^ Mendoza H, Perlin MH, Schirawski J (May 2020). "Mitochondrial Inheritance in Phytopathogenic Fungi-Everything Is Known, or Is It?". Int J Mol Sci. 21 (11): 3883. doi:10.3390/ijms21113883. PMC 7312866. PMID 32485941.
- ^ a b Luo S, Valencia CA, Zhang J, Lee NC, Slone J, Gui B, et al. (December 2018). "Biparental Inheritance of Mitochondrial DNA in Humans". Proceedings of the National Academy of Sciences of the United States of America. 115 (51): 13039–13044. Bibcode:2018PNAS..11513039L. doi:10.1073/pnas.1810946115. PMC 6304937. PMID 30478036.
- ^ Pagnamenta AT, Wei W, Rahman S, Chinnery PF (August 2021). "Biparental inheritance of mitochondrial DNA revisited". Nature Reviews. Genetics. 22 (8): 477–478. doi:10.1038/s41576-021-00380-6. PMID 34031572. S2CID 235199218.
- ^ Salas A, Schönherr S, Bandelt HJ, Gómez-Carballa A, Weissensteiner H (July 2020). "Extraordinary claims require extraordinary evidence in asserted mtDNA biparental inheritance". Forensic Science International. Genetics. 47: 102274. doi:10.1016/j.fsigen.2020.102274. PMID 32330850. S2CID 216131636.
- ^ Wei W, Pagnamenta AT, Gleadall N, Sanchis-Juan A, Stephens J, Broxholme J, Tuna S, Odhams CA, Fratter C, Turro E, Caulfield MJ, Taylor JC, Rahman S, Chinnery PF (April 2020). "Nuclear-mitochondrial DNA segments resemble paternally inherited mitochondrial DNA in humans". Nature Communications. 11 (1): 1740. Bibcode:2020NatCo..11.1740W. doi:10.1038/s41467-020-15336-3. PMC 7142097. PMID 32269217.
- ^ Sutovsky P, Moreno RD, Ramalho-Santos J, Dominko T, Simerly C, Schatten G (November 1999). "Ubiquitin tag for sperm mitochondria". Nature. 402 (6760): 371–372. Bibcode:1999Natur.402..371S. doi:10.1038/46466. PMID 10586873. S2CID 205054671. Discussed in: Travis J (2000). "Mom's Eggs Execute Dad's Mitochondria". Science News. 157 (1): 5. doi:10.2307/4012086. JSTOR 4012086. Archived from the original on 19 December 2007.
- ^ "Hiring a DNA Testing Company Genealogy". Family Search. The Church of Jesus Christ of Latter-day Saints. 2016. Archived from the original on 3 October 2016. Retrieved 2 October 2016.
- ^ Vilà C, Savolainen P, Maldonado JE, Amorim IR, Rice JE, Honeycutt RL, et al. (June 1997). "Multiple and ancient origins of the domestic dog" (PDF). Science. 276 (5319): 1687–1689. doi:10.1126/science.276.5319.1687. PMID 9180076. Archived (PDF) from the original on 9 October 2022.
- ^ Wolff JN, White DJ, Woodhams M, White HE, Gemmell NJ (2011). "The strength and timing of the mitochondrial bottleneck in salmon suggests a conserved mechanism in vertebrates". PLOS ONE. 6 (5): e20522. Bibcode:2011PLoSO...620522W. doi:10.1371/journal.pone.0020522. PMC 3105079. PMID 21655224.
- ^ Cree LM, Samuels DC, de Sousa Lopes SC, Rajasimha HK, Wonnapinij P, Mann JR, et al. (February 2008). "A reduction of mitochondrial DNA molecules during embryogenesis explains the rapid segregation of genotypes". Nature Genetics. 40 (2): 249–254. doi:10.1038/ng.2007.63. PMID 18223651. S2CID 205344980.
- ^ Cao L, Shitara H, Horii T, Nagao Y, Imai H, Abe K, et al. (March 2007). "The mitochondrial bottleneck occurs without reduction of mtDNA content in female mouse germ cells". Nature Genetics. 39 (3): 386–390. doi:10.1038/ng1970. PMID 17293866. S2CID 10686347.
- ^ Wai T, Teoli D, Shoubridge EA (December 2008). "The mitochondrial DNA genetic bottleneck results from replication of a subpopulation of genomes". Nature Genetics. 40 (12): 1484–1488. doi:10.1038/ng.258. PMID 19029901. S2CID 225349.
- ^ Alexander M, Ho SY, Molak M, Barnett R, Carlborg Ö, Dorshorst B, et al. (October 2015). "Mitogenomic analysis of a 50-generation chicken pedigree reveals a rapid rate of mitochondrial evolution and evidence for paternal mtDNA inheritance". Biology Letters. 11 (10): 20150561. doi:10.1098/rsbl.2015.0561. PMC 4650172. PMID 26510672.
- ^ Gyllensten U, Wharton D, Josefsson A, Wilson AC (July 1991). "Paternal inheritance of mitochondrial DNA in mice". Nature. 352 (6332): 255–257. Bibcode:1991Natur.352..255G. doi:10.1038/352255a0. PMID 1857422. S2CID 4278149.
- ^ Shitara H, Hayashi JI, Takahama S, Kaneda H, Yonekawa H (February 1998). "Maternal inheritance of mouse mtDNA in interspecific hybrids: segregation of the leaked paternal mtDNA followed by the prevention of subsequent paternal leakage". Genetics. 148 (2): 851–857. doi:10.1093/genetics/148.2.851. PMC 1459812. PMID 9504930.
- ^ Zhao X, Li N, Guo W, Hu X, Liu Z, Gong G, et al. (October 2004). "Further evidence for paternal inheritance of mitochondrial DNA in the sheep (Ovis aries)". Heredity. 93 (4): 399–403. doi:10.1038/sj.hdy.6800516. PMID 15266295.
- ^ Steinborn R, Zakhartchenko V, Jelyazkov J, Klein D, Wolf E, Müller M, Brem G (April 1998). "Composition of parental mitochondrial DNA in cloned bovine embryos". FEBS Letters. 426 (3): 352–356. doi:10.1016/S0014-5793(98)00350-0. PMID 9600265.
- ^ Singh AN (April 2012). "A105 Family Decoded: Discovery of Genome-Wide Fingerprints for Personalized Genomic Medicine". ScienceMED. 3 (2): 115–126.
- ^ Singh AN (May 2018). "Customized Biomedical Informatics". BMC Big Data Analytics. 3. doi:10.1186/s41044-018-0030-3.
- ^ Schwartz M, Vissing J (August 2002). "Paternal inheritance of mitochondrial DNA". The New England Journal of Medicine. 347 (8): 576–580. doi:10.1056/NEJMoa020350. PMID 12192017.
- ^ Passamonti M, Ghiselli F (February 2009). "Doubly uniparental inheritance: two mitochondrial genomes, one precious model for organelle DNA inheritance and evolution". DNA and Cell Biology. 28 (2): 79–89. doi:10.1089/dna.2008.0807. PMID 19196051.
- ^ Kondo R, Matsuura ET, Chigusa SI (April 1992). "Further observation of paternal transmission of Drosophila mitochondrial DNA by PCR selective amplification method". Genetical Research. 59 (2): 81–84. doi:10.1017/S0016672300030287. PMID 1628820.
- ^ Wolff JN, Nafisinia M, Sutovsky P, Ballard JW (January 2013). "Paternal transmission of mitochondrial DNA as an integral part of mitochondrial inheritance in metapopulations of Drosophila simulans". Heredity. 110 (1): 57–62. doi:10.1038/hdy.2012.60. PMC 3522233. PMID 23010820.
- ^ Meusel MS, Moritz RF (December 1993). "Transfer of paternal mitochondrial DNA during fertilization of honeybee (Apis mellifera L.) eggs". Current Genetics. 24 (6): 539–543. doi:10.1007/BF00351719. PMID 8299176. S2CID 15621371.
- ^ Fontaine KM, Cooley JR, Simon C (September 2007). "Evidence for paternal leakage in hybrid periodical cicadas (Hemiptera: Magicicada spp.)". PLOS ONE. 2 (9): e892. Bibcode:2007PLoSO...2..892F. doi:10.1371/journal.pone.0000892. PMC 1963320. PMID 17849021.
- ^ Frith M (14 October 2003). "Ban on scientists trying to create three-parent baby". The Independent.[permanent dead link]
- ^ Roberts M (27 September 2016). "First 'three person baby' born using new method". BBC News. Archived from the original on 30 May 2019. Retrieved 28 September 2016.
- ^ a b Alexeyev MF (October 2009). "Is there more to aging than mitochondrial DNA and reactive oxygen species?". The FEBS Journal. 276 (20): 5768–5787. doi:10.1111/j.1742-4658.2009.07269.x. PMC 3097520. PMID 19796285.
- ^ Anson RM, Hudson E, Bohr VA (February 2000). "Mitochondrial endogenous oxidative damage has been overestimated". FASEB Journal. 14 (2): 355–360. doi:10.1096/fasebj.14.2.355. PMID 10657991. S2CID 19771110.
- ^ Thorslund T, Sunesen M, Bohr VA, Stevnsner T (April 2002). "Repair of 8-oxoG is slower in endogenous nuclear genes than in mitochondrial DNA and is without strand bias". DNA Repair. 1 (4): 261–273. doi:10.1016/S1568-7864(02)00003-4. PMID 12509245. Archived from the original on 31 December 2019. Retrieved 30 June 2019.
- ^ Guliaeva NA, Kuznetsova EA, Gaziev AI (2006). "[Proteins associated with mitochondrial DNA protect it against the action of X-rays and hydrogen peroxide]" [Proteins associated with mitochondrial DNA protect it against the action of X-rays and hydrogen peroxide]. Biofizika (in Russian). 51 (4): 692–697. PMID 16909848.
- ^ Alexeyev M, Shokolenko I, Wilson G, LeDoux S (May 2013). "The maintenance of mitochondrial DNA integrity--critical analysis and update". Cold Spring Harbor Perspectives in Biology. 5 (5): a012641. doi:10.1101/cshperspect.a012641. PMC 3632056. PMID 23637283.
- ^ Hogan CM (2010). "Mutation". In Monosson E, Cleveland CJ (eds.). Encyclopedia of Earth. Washington DC: National Council for Science and the Environment. Archived from the original on 30 April 2011. Retrieved 18 April 2011.
- ^ Alexeyev MF, Ledoux SP, Wilson GL (October 2004). "Mitochondrial DNA and aging". Clinical Science. 107 (4): 355–364. doi:10.1042/CS20040148. PMID 15279618. S2CID 5747202.
- ^ Burgstaller JP, Johnston IG, Poulton J (January 2015). "Mitochondrial DNA disease and developmental implications for reproductive strategies". Molecular Human Reproduction. 21 (1): 11–22. doi:10.1093/molehr/gau090. PMC 4275042. PMID 25425607.
- ^ Burgstaller JP, Johnston IG, Jones NS, Albrechtová J, Kolbe T, Vogl C, et al. (June 2014). "MtDNA segregation in heteroplasmic tissues is common in vivo and modulated by haplotype differences and developmental stage". Cell Reports. 7 (6): 2031–2041. doi:10.1016/j.celrep.2014.05.020. PMC 4570183. PMID 24910436.
- ^ a b Taylor RW, Turnbull DM (May 2005). "Mitochondrial DNA mutations in human disease". Nature Reviews. Genetics. 6 (5): 389–402. doi:10.1038/nrg1606. PMC 1762815. PMID 15861210.
- ^ Angelini C, Bello L, Spinazzi M, Ferrati C (July 2009). "Mitochondrial disorders of the nuclear genome". Acta Myologica. 28 (1): 16–23. PMC 2859630. PMID 19772191.
- ^ Reguly B, Jakupciak JP, Parr RL (October 2010). "3.4 kb mitochondrial genome deletion serves as a surrogate predictive biomarker for prostate cancer in histopathologically benign biopsy cores". Canadian Urological Association Journal. 4 (5): E118–E122. doi:10.5489/cuaj.932. PMC 2950771. PMID 20944788.
- ^ Robinson K, Creed J, Reguly B, Powell C, Wittock R, Klein D, et al. (June 2010). "Accurate prediction of repeat prostate biopsy outcomes by a mitochondrial DNA deletion assay". Prostate Cancer and Prostatic Diseases. 13 (2): 126–131. doi:10.1038/pcan.2009.64. PMID 20084081. S2CID 25050759.
- ^ Mair R, Mouliere F, Smith CG, Chandrananda D, Gale D, Marass F, et al. (January 2019). "Measurement of Plasma Cell-Free Mitochondrial Tumor DNA Improves Detection of Glioblastoma in Patient-Derived Orthotopic Xenograft Models". Cancer Research. 79 (1): 220–230. doi:10.1158/0008-5472.CAN-18-0074. PMC 6753020. PMID 30389699. Archived from the original on 24 September 2019. Retrieved 24 September 2019.
- ^ de Grey A (1999). The Mitochondrial Free Radical Theory of Aging (PDF). Landes Bioscience. ISBN 978-1-57059-564-6. Archived (PDF) from the original on 3 June 2016. Retrieved 1 May 2016.[page needed]
- ^ Lewis KN, Andziak B, Yang T, Buffenstein R (October 2013). "The naked mole-rat response to oxidative stress: just deal with it". Antioxidants & Redox Signaling. 19 (12): 1388–1399. doi:10.1089/ars.2012.4911. PMC 3791056. PMID 23025341.
- ^ Shigenaga MK, Hagen TM, Ames BN (November 1994). "Oxidative damage and mitochondrial decay in aging". Proceedings of the National Academy of Sciences of the United States of America. 91 (23): 10771–10778. Bibcode:1994PNAS...9110771S. doi:10.1073/pnas.91.23.10771. JSTOR 2365473. PMC 45108. PMID 7971961.
- ^ Trifunovic A, Hansson A, Wredenberg A, Rovio AT, Dufour E, Khvorostov I, et al. (December 2005). "Somatic mtDNA mutations cause aging phenotypes without affecting reactive oxygen species production". Proceedings of the National Academy of Sciences of the United States of America. 102 (50): 17993–17998. Bibcode:2005PNAS..10217993T. doi:10.1073/pnas.0508886102. JSTOR 4152716. PMC 1312403. PMID 16332961.
- ^ Aledo JC, Li Y, de Magalhães JP, Ruíz-Camacho M, Pérez-Claros JA (April 2011). "Mitochondrially encoded methionine is inversely related to longevity in mammals". Aging Cell. 10 (2): 198–207. doi:10.1111/j.1474-9726.2010.00657.x. PMID 21108730.
- ^ Shabalina IG, Vyssokikh MY, Gibanova N, Csikasz RI, Edgar D, Hallden-Waldemarson A, et al. (February 2017). "Improved health-span and lifespan in mtDNA mutator mice treated with the mitochondrially targeted antioxidant SkQ1". Aging. 9 (2): 315–339. doi:10.18632/aging.101174. PMC 5361666. PMID 28209927.
- ^ Ferrari CK (2004). "Functional foods, herbs and nutraceuticals: towards biochemical mechanisms of healthy aging". Biogerontology. 5 (5): 275–289. doi:10.1007/s10522-004-2566-z. PMID 15547316. S2CID 11568208.
- ^ Taylor RW (February 2005). "Gene therapy for the treatment of mitochondrial DNA disorders". Expert Opinion on Biological Therapy. 5 (2): 183–194. doi:10.1517/14712598.5.2.183. PMID 15757380. S2CID 35276183.
- ^ Bjelakovic G, Nikolova D, Gluud C (September 2013). "Antioxidant supplements to prevent mortality". JAMA. 310 (11): 1178–1179. doi:10.1001/jama.2013.277028. PMID 24045742.
- ^ Gureev AP, Andrianova NV, Pevzner IB, Zorova LD, Chernyshova EV, Sadovnikova IS, et al. (September 2022). "Dietary restriction modulates mitochondrial DNA damage and oxylipin profile in aged rats". The FEBS Journal. 289 (18): 5697–5713. doi:10.1111/febs.16451. PMID 35373508. S2CID 247938550. Archived from the original on 4 May 2022. Retrieved 11 May 2022.
- ^ Wang J, Xiong S, Xie C, Markesbery WR, Lovell MA (May 2005). "Increased oxidative damage in nuclear and mitochondrial DNA in Alzheimer's disease". Journal of Neurochemistry. 93 (4): 953–962. doi:10.1111/j.1471-4159.2005.03053.x. PMID 15857398.
- ^ Bonda DJ, Wang X, Lee HG, Smith MA, Perry G, Zhu X (April 2014). "Neuronal failure in Alzheimer's disease: a view through the oxidative stress looking-glass". Neuroscience Bulletin. 30 (2): 243–252. doi:10.1007/s12264-013-1424-x. PMC 4097013. PMID 24733654.
- ^ Canugovi C, Shamanna RA, Croteau DL, Bohr VA (June 2014). "Base excision DNA repair levels in mitochondrial lysates of Alzheimer's disease". Neurobiology of Aging. 35 (6): 1293–1300. doi:10.1016/j.neurobiolaging.2014.01.004. PMC 5576885. PMID 24485507.
- ^ Liu Z, Zhou T, Ziegler AC, Dimitrion P, Zuo L (2017). "Oxidative Stress in Neurodegenerative Diseases: From Molecular Mechanisms to Clinical Applications". Oxidative Medicine and Cellular Longevity. 2017: 2525967. doi:10.1155/2017/2525967. PMC 5529664. PMID 28785371.
- ^ Ayala-Peña S (September 2013). "Role of oxidative DNA damage in mitochondrial dysfunction and Huntington's disease pathogenesis". Free Radical Biology & Medicine. 62: 102–110. doi:10.1016/j.freeradbiomed.2013.04.017. PMC 3722255. PMID 23602907.
- ^ Kikuchi H, Furuta A, Nishioka K, Suzuki SO, Nakabeppu Y, Iwaki T (April 2002). "Impairment of mitochondrial DNA repair enzymes against accumulation of 8-oxo-guanine in the spinal motor neurons of amyotrophic lateral sclerosis". Acta Neuropathologica. 103 (4): 408–414. doi:10.1007/s00401-001-0480-x. PMID 11904761. S2CID 2102463.
- ^ Lehmann G, Budovsky A, Muradian KK, Fraifeld VE (2006). "Mitochondrial genome anatomy and species-specific lifespan". Rejuvenation Research. 9 (2): 223–226. doi:10.1089/rej.2006.9.223. PMID 16706648.
- ^ a b Lehmann G, Segal E, Muradian KK, Fraifeld VE (April 2008). "Do mitochondrial DNA and metabolic rate complement each other in determination of the mammalian maximum longevity?". Rejuvenation Research. 11 (2): 409–417. doi:10.1089/rej.2008.0676. PMID 18442324.
- ^ Lehmann G, Muradian KK, Fraifeld VE (2013). "Telomere length and body temperature-independent determinants of mammalian longevity?". Frontiers in Genetics. 4 (111): 111. doi:10.3389/fgene.2013.00111. PMC 3680702. PMID 23781235.
- ^ Toren D, Barzilay T, Tacutu R, Lehmann G, Muradian KK, Fraifeld VE (January 2016). "MitoAge: a database for comparative analysis of mitochondrial DNA, with a special focus on animal longevity". Nucleic Acids Research. 44 (D1): D1262–D1265. doi:10.1093/nar/gkv1187. PMC 4702847. PMID 26590258.
- ^ Mikhailova AG, Mikhailova AA, Ushakova K, Tretiakov EO, Iliushchenko D, Shamansky V, et al. (October 2022). "A mitochondria-specific mutational signature of aging: increased rate of A > G substitutions on the heavy strand". Nucleic Acids Research. 50 (18): 10264–10277. doi:10.1093/nar/gkac779. PMC 9561281. PMID 36130228.
- ^ Damas J, Carneiro J, Gonçalves J, Stewart JB, Samuels DC, Amorim A, Pereira F (September 2012). "Mitochondrial DNA deletions are associated with non-B DNA conformations". Nucleic Acids Research. 40 (16): 7606–7621. doi:10.1093/nar/gks500. PMC 3439893. PMID 22661583.
- ^ Oliveira PH, da Silva CL, Cabral JM (2013). "An appraisal of human mitochondrial DNA instability: new insights into the role of non-canonical DNA structures and sequence motifs". PLOS ONE. 8 (3): e59907. Bibcode:2013PLoSO...859907O. doi:10.1371/journal.pone.0059907. PMC 3612095. PMID 23555828.
- ^ Brown WM, George M, Wilson AC (April 1979). "Rapid evolution of animal mitochondrial DNA". Proceedings of the National Academy of Sciences of the United States of America. 76 (4): 1967–1971. Bibcode:1979PNAS...76.1967B. doi:10.1073/pnas.76.4.1967. JSTOR 69636. PMC 383514. PMID 109836.
- ^ Butler JM (2005). Forensic DNA typing : biology, technology, and genetics of STR markers (2nd ed.). Amsterdam: Elsevier Academic Press. pp. 241–242, 246, 258–259. ISBN 0-12-147952-8.
- ^ Syndercombe Court D (September 2021). "Mitochondrial DNA in forensic use". Emerging Topics in Life Sciences. 5 (3): 415–426. doi:10.1042/ETLS20210204. PMC 8457767. PMID 34374411.
- ^ Davis CL (1998). "Mitochondrial DNA: State of Tennessee v. Paul Ware" (PDF). Profiles in DNA. 1 (3): 6–7. Archived (PDF) from the original on 21 August 2016. Retrieved 21 October 2013.
- ^ [1] Archived 21 August 2016 at the Wayback Machine Court case name listed in the appeal.[full citation needed] Retrieved 17 April 2015.
- ^ [2] Archived 17 May 2015 at the Wayback Machine[self-published source?] Defense lawyer. Retrieved 17 April 2015.
- ^ Garlicki D (11 March 1998). "DNA Tests Got Rorrer Life in Jail". The Morning Call.
- ^ Pellegrino P, Thomas P (2000), "Forensic files. a woman scorned Episode 55 Episode 55", Courtroom Television Network, Medstar Television, OCLC 456260156
- ^ "Judge allows DNA in Samantha Runnion case". Associated Press. 18 February 2005. Retrieved 4 April 2007.
- ^ "Canine DNA Admitted In California Murder Case". Pit Bulletin Legal News. 5 December 2013. Archived from the original on 2 February 2014. Retrieved 21 January 2014.
- ^ Kennedy M (4 February 2013). "Richard III: DNA confirms twisted bones belong to king". The Guardian. Archived from the original on 8 January 2019. Retrieved 7 December 2014.
- ^ Xu X, Arnason U (May 1996). "A complete sequence of the mitochondrial genome of the western lowland gorilla". Molecular Biology and Evolution. 13 (5): 691–698. doi:10.1093/oxfordjournals.molbev.a025630. PMID 8676744. Archived from the original on 4 August 2020. Retrieved 3 February 2020.
- ^ Wei W, Schon KR, Elgar G, Orioli A, Tanguy M, Giess A, et al. (November 2022). "Nuclear-embedded mitochondrial DNA sequences in 66,083 human genomes". Nature. 611 (7934): 105–114. Bibcode:2022Natur.611..105W. doi:10.1038/s41586-022-05288-7. PMC 9630118. PMID 36198798.
- ^ Wei W, Schon KR, Elgar G, Orioli A, Tanguy M, Giess A, et al. (November 2022). "Nuclear-embedded mitochondrial DNA sequences in 66,083 human genomes". Nature. 611 (7934): 105–114. Bibcode:2022Natur.611..105W. doi:10.1038/s41586-022-05288-7. PMC 9630118. PMID 36198798.
- ^ Nass MM, Nass S (December 1963). "Intramitochondrial Fibers with DNA Characteristics". The Journal of Cell Biology. 19 (3): 593–611. doi:10.1083/jcb.19.3.593. PMC 2106331. PMID 14086138.
- ^ Schatz G, Haslbrunner E, Tuppy H (March 1964). "Deoxyribonucleic acid associated with yeast mitochondria". Biochemical and Biophysical Research Communications. 15 (2): 127–132. doi:10.1016/0006-291X(64)90311-0. PMID 26410904.
- ^ Ehler E, Novotný J, Juras A, Chylenski M, Moravcík O, Paces J (January 2019). "AmtDB: a database of ancient human mitochondrial genomes". Nucleic Acids Research. 47 (D1): D29–D32. doi:10.1093/nar/gky843. PMC 6324066. PMID 30247677.
- ^ Gu Z, Li J, Gao S, Gong M, Wang J, Xu H, et al. (June 2011). "InterMitoBase: an annotated database and analysis platform of protein-protein interactions for human mitochondria". BMC Genomics. 12: 335. doi:10.1186/1471-2164-12-335. PMC 3142533. PMID 21718467.
- ^ Damas J, Carneiro J, Amorim A, Pereira F (January 2014). "MitoBreak: the mitochondrial DNA breakpoints database". Nucleic Acids Research. 42 (Database issue): D1261–D1268. doi:10.1093/nar/gkt982. PMC 3965124. PMID 24170808.
- ^ Iwasaki W, Fukunaga T, Isagozawa R, Yamada K, Maeda Y, Satoh TP, et al. (November 2013). "MitoFish and MitoAnnotator: a mitochondrial genome database of fish with an accurate and automatic annotation pipeline". Molecular Biology and Evolution. 30 (11): 2531–2540. doi:10.1093/molbev/mst141. PMC 3808866. PMID 23955518.
- ^ Cawthorn DM, Steinman HA, Witthuhn RC (November 2011). "Establishment of a mitochondrial DNA sequence database for the identification of fish species commercially available in South Africa". Molecular Ecology Resources. 11 (6): 979–991. doi:10.1111/j.1755-0998.2011.03039.x. PMID 21689383. S2CID 205971257.
- ^ Lee YS, Oh J, Kim YU, Kim N, Yang S, Hwang UW (January 2008). "Mitome: dynamic and interactive database for comparative mitochondrial genomics in metazoan animals". Nucleic Acids Research. 36 (Database issue): D938–D942. doi:10.1093/nar/gkm763. PMC 2238945. PMID 17940090.
- ^ Catalano D, Licciulli F, Turi A, Grillo G, Saccone C, D'Elia D (January 2006). "MitoRes: a resource of nuclear-encoded mitochondrial genes and their products in Metazoa". BMC Bioinformatics. 7: 36. doi:10.1186/1471-2105-7-36. PMC 1395343. PMID 16433928.
- ^ Kumar M, Kapil A, Shanker A (November 2014). "MitoSatPlant: mitochondrial microsatellites database of viridiplantae". Mitochondrion. 19 (Pt B): 334–337. doi:10.1016/j.mito.2014.02.002. PMID 24561221.
- ^ D'Onorio de Meo P, D'Antonio M, Griggio F, Lupi R, Borsani M, Pavesi G, et al. (January 2012). "MitoZoa 2.0: a database resource and search tools for comparative and evolutionary analyses of mitochondrial genomes in Metazoa". Nucleic Acids Research. 40 (Database issue): D1168–D1172. doi:10.1093/nar/gkr1144. PMC 3245153. PMID 22123747.
- ^ "Mothers can influence offspring's height, lifespan and disease risk through mitochondria". phys.org. Archived from the original on 14 June 2021. Retrieved 14 June 2021.
- ^ Yonova-Doing E, Calabrese C, Gomez-Duran A, Schon K, Wei W, Karthikeyan S, et al. (July 2021). "An atlas of mitochondrial DNA genotype-phenotype associations in the UK Biobank". Nature Genetics. 53 (7): 982–993. doi:10.1038/s41588-021-00868-1. PMC 7611844. PMID 34002094. S2CID 234768578.
External links
 Media related to Mitochondrial DNA at Wikimedia Commons
Media related to Mitochondrial DNA at Wikimedia Commons
Types of nucleic acids | |||||||
|---|---|---|---|---|---|---|---|
| Constituents | |||||||
| Ribonucleic acids (coding, non-coding) |
| ||||||
| Deoxyribonucleic acids | |||||||
| Analogues | |||||||
| Cloning vectors | |||||||