
| Homo ("humans") Temporal range: Piacenzian-Present,
| |
|---|---|
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Primates |
| Suborder: | Haplorhini |
| Infraorder: | Simiiformes |
| Family: | Hominidae |
| Subfamily: | Homininae |
| Tribe: | Hominini |
| Genus: | Homo Linnaeus, 1758 |
| Type species | |
| Homo sapiens Linnaeus, 1758
| |
| Species | |
other species or subspecies suggested | |
| Synonyms | |
|
Synonyms
| |
−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — |
| |||||||||||||||||||||||||||||
Human taxonomy is the classification of the human species (systematic name Homo sapiens, Latin: "wise man") within zoological taxonomy. The systematic genus, Homo, is designed to include both anatomically modern humans and extinct varieties of archaic humans. Current humans have been designated as subspecies Homo sapiens sapiens, differentiated, according to some, from the direct ancestor, Homo sapiens idaltu (with some other research instead classifying idaltu and current humans as belonging to the same subspecies[1][2][3]).
Since the introduction of systematic names in the 18th century, knowledge of human evolution has increased drastically, and a number of intermediate taxa have been proposed in the 20th and early 21st centuries. The most widely accepted taxonomy grouping takes the genus Homo as originating between two and three million years ago, divided into at least two species, archaic Homo erectus and modern Homo sapiens, with about a dozen further suggestions for species without universal recognition.
The genus Homo is placed in the tribe Hominini alongside Pan (chimpanzees). The two genera are estimated to have diverged over an extended time of hybridization, spanning roughly 10 to 6 million years ago, with possible admixture as late as 4 million years ago. A subtribe of uncertain validity, grouping archaic "pre-human" or "para-human" species younger than the Homo-Pan split, is Australopithecina (proposed in 1939).
A proposal by Wood and Richmond (2000) would introduce Hominina as a subtribe alongside Australopithecina, with Homo the only known genus within Hominina. Alternatively, following Cela-Conde and Ayala (2003), the "pre-human" or "proto-human" genera of Australopithecus, Ardipithecus, Praeanthropus, and possibly Sahelanthropus, may be placed on equal footing alongside the genus Homo. An even more extreme view rejects the division of Pan and Homo as separate genera, which based on the Principle of Priority would imply the reclassification of chimpanzees as Homo paniscus (or similar).[4]
Categorizing humans based on phenotypes is a socially controversial subject. Biologists originally classified races as subspecies, but contemporary anthropologists reject the concept of race as a useful tool to understanding humanity, and instead view humanity as a complex, interrelated genetic continuum. Taxonomy of the hominins continues to evolve.[5][6]
YouTube Encyclopedic
-
1/5Views:3 134 565187 18719 3834 8801 687
-
Taxonomy: Life's Filing System - Crash Course Biology #19
-
Taxonomy and Taxonomic Hierarchy (Biological Classification of Living Things)
-
WACE Human Biology: Human Classification
-
Human Classification
-
Taxonomy and The Tree of Life | Human Taxonomic Ranking | Bio 101 | STEMstream
Transcription
Taxonomy! It's the science of classifying living things. That sounds exciting. Today we'll basically be learning the Dewey Decimel System of evolution! It's like filing! You must be on the edge of your seat. OK, shut up. When it comes down to it, this science doesn't just categorize organisms, when you look a little deeper, you realize it's telling the story of all life on earth. And it's a pretty good story. Every living thing on this planet is related to every other living thing. If you go far enough back, we all have a common ancestor. An organism that both you and I are descended from. Or something that a star fish and a blue whale are descended from. Or, even weirder, that an oak tree and a salmon are both descended from. That organism lived. It lived very long ago. But it was here. And I dig that. The trick of taxonomy, is basically figuring out where all those branches of the evolutionary tree are, and finding some convenient labels to help us understand all of these remarkable interrelationships. Let's be clear though, taxonomy isn't about describing life in all of it's ridiculous detail, it's mostly about helping humans understand it, because it's way too complicated without structure. To get that structure biologists use the taxonomic system to classify all the organisms on the Earth. It's sometimes called the Phylogenetic Tree, or the Tree of Life, and it illustrates the evolutionary relationships between all living species. There are about 2 million known species, but there could be anywhere from 5 million to 100 million species scientists really have no freaking idea. New species keep getting discovered all the time, and the more organisms we have to keep track of, the more complex the Phylogenetic Tree becomes. So, there's not always a consensus about how to classify this stuff. There's a lot of gray area in the Natural World. Actually, let me rephrase that: the Natural World is one giant Gray Area. Sometimes it's just hard to know where to put a certain group of organisms, and eventually the group gets so big, the classification system has to be messed with to make room for it. So, the system isn't perfect, but it's good enough that we've been using it for around 250 years. [Sniffing] What's that? Do you smell a Bio-lography coming on? Carl Linnaeus was a Swede born in 1707. And early in his career as a botanist he realized that the botanical nomenclature of 18th century Europe was.. well, just crap. For instance, in his day, the "formal" name of a tomato plant was Solanum caule inerme herbaceo, foliis pinnatis incisis, racemis simplicibus. Linnaeus actually said once, "I shudder at the sight of most botanical names given by modern authorities." Not only did this sloppiness bother him, he saw a whole sugarstorm blowing in: New plants were still being discovered in Europe, but that was nothing compared to the crazy stuff that was coming from the New World. Linnaeus saw that pretty soon, naming conventions were just going to collapse under all these new things to name. And THEN what? Linnaeus famously started off by naming himself. He came from a peasant family, and at the time, surnames were just for rich people, so when Carl went to college, they asked him for his surname and he just made one up: Linneaus, after the Linden trees that grew on his family's homestead. Linnaeus got a medical degree and became a professor at Uppsala University where he devoted himself to the study of nomenclature. He had his students go places and bring back specimens for him to study and categorize. The method he eventually adopted was based on morphology, or physical form and structure. This wasn't necessarily a new idea. Back then, people grouped organisms by analogous or homoplasic traits, structures that appear similar but actually come from completely independent origins. By this definition, birds would be more closely related to butterflies than to reptiles because birds and butterflies can both fly. But Linnaeus had a good mind for this stuff and turned out to have a real knack for choosing actual homologous traits for his classification system traits that stem from a common evolutionary ancestor. Linnaeus didn't know jack about evolution Darwin wouldn't come around for another 100 plus years but he just intuited that some traits were more important than others. For instance, he was struck by the fact that reproductive apparatus seemed to be a good way of classifying plants. He also caused a scandal by classifying the Class Mammalia based on the female's ability to produce milk from their nipples. Because apparently that was pretty racy stuff back then. In his lifetime Linnaeus catalogued roughly 7,700 plants and 4,400 animals, and he published his classifications in a catalog called Systema Naturae, which by the time he wrote the 12th edition, was 2,300 pages long. In the meantime, Linneaus actually adopted a personal motto: "God created, Linnaeus organized." Although taxonomy has come a long way since Linnaeus, we still use a bunch of the conventions that he invented. For instance, we still arrange things into taxa, or groups of organisms, and we still us the same Taxa as Linnaeus: kingdom, phylum, class, order, family, genus and species. We also still use Linnaeus' convention of binomial nomenclature using a unique, two-part name for every species the genus and species name, in Latin or Latin-ish. This practice actually started back in the Middle Ages when educated people were expected to know Latin. We know a lot less latin now, but we know a lot more about evolution which Linnaeus didn't. And we have technologies like genetic testing to classify relationships between organisms. And yet we still use Linnaeus's morphology-based system because genetic evidence generally agrees with classifications that are made based on structure and form. However, because there was a lot of life that Linnaeus had no idea about, we had to stick a new taxa above Linnaeus' Kingdom. We call it Domain. And it's as broad as you can get. The Domains are Bacteria, Archaea and Eukarya. The bacteria and archae are prokaryotes, meaning their genetic material goes commando with no nucleus to enclose it. While the Eukarya make up all the life forms with a nucleus and include pretty much all the life that you think of as life, and quite a lot of the life that you don't think about at all. It might seem like, since all macroscopic life only gets one domain, it's kinda silly to give prokaryotes two and for a long time, we didn't. We didn't divide them up into different domains. They hung out together in a single domain called Monera. But it later became clear that Bacteria, which live pretty much everywhere on earth, including inside of you and deep in the Earth's crust, and Archaea which are even more hardy than bacteria, have distinct evolutionary histories. Archaea being more closely related to eukaryotes and, yes, thus me and you. They have totally different cell membranes and the enzymes they use to make RNA, their RNA polymerase, is much more like ours. Under the domain Eukarya, which is by far the most interesting and even occasionally adorable domain, we have Kingdoms: Protista, Fungi, Plantae and Animalia. Now, scientists have settled on these four. For now. But these are categories that are a human creation, but there are good reasons for that human creation. The unscientific truth is that we looked at life and divided it up based on what we saw. So we were like, "Well, protists are single-celled organisms, so, they're very different from the rest of the domain. Plants get their energy from the sun and fungi look and act very different from plants and animals, and we already know what animals are, so they have to get their own kingdom." And though scientists are loathe to admit it, that system of just looking and dividing things up actually worked pretty well for us. Not perfectly, but pretty well. But there's a reason why this worked so well. Evolutionarily, there are actual categories. Each of these kingdoms is a huge branch in the tree of life. At each branch, an evolutionary change occurred that was so massively helpful that it spawned a vast diversity of descendents. Plants or Plantae are the autotrophs of the Domain Eukarya. Autotrophs meaning that they can feed themselves, through photosynthesis of course. Their cellulose-based cell walls and chloroplasts giving them a distinct difference from all other multi-cellular life. There are two other sorts of -trophs. The heterotrophs, which get their energy by eating other organisms. And Chemotrophs, which are weird and crazy and only show up in bacteria and achaea, and they get their energy from chemicals. Now the kingdom Protista is weird because it contains both autotrophs and heterotrophs. Some protists can photosynthesize, while others eat living things. Protists are basically a bunch of weird, eukaryotic single-celled organisms that may or may not be evolutionarily related to each other scientists are still trying to figure it out. Some are plant-like, like algae, some are more animal-like, like amoebas, and some are fungus-like, like slime molds. Protists are one of those gray areas I was telling you about. So don't be surprised if, by the time you're teaching this to your biology students, there are more than four kingdoms in Eukarya. Fungi, which are, you know—the funguses. They include mushrooms, smuts, puffballs, truffles, molds, and yeasts and they're pretty cool because they have cell walls like plants, but instead of being made of cellulose, they're made of another carbohydrate called chitin, which is also what the beak of a giant squid is made out of, or the exoskeleton of a beetle. Because fungi are heterotrophs like animals, they have these sort of digestive enzymes that break down their food and get reabsorbed. But they can't move, they don't require a stomach for digestion they just grow on top of whatever it is they're digesting and digest it right where it is. Which is super convenient! And finally, we have Kingdom Animalia. Which is the lovely kingdom that we find ourselves and 100% of adorable organisms in. Animals are multicellular, always. We're heterotrophic, so we spend a lot of our time hunting down food because we can't make it ourselves. Almost all of us can move, at least during some stage of our life cycle. And most of us develop either two or three germ layers during embryonic development, wait for it... ...unless you're a sponge. So like I said, we use this taxonomic system to describe the common ancestry and evolutionary history of an organism. Looking at the phylogenetic tree, you can tell that humans are more closely related to mice than we are to fish, and more closely related to fish than we are to fruit flies. So how about we pick an organism and follow it all the way through the taxa, from kingdom to species, just to see how it works. I know! Let's pick this kitty. Because I know she'd like it. Right, cat? So, kitties have cells that have nuclei and membrane surrounded organelles. And they're multicellular and heterotrophic and have three germ layers of cells when they're embryos, so they're in the kingdom Animalia. And they have a spinal cord running down their backs, protected by vertebrae, and disks in between them. And they have a tail that doesn't have a butthole at the end of it like a worm, which I'm really glad about. And that puts her in the phylum Chordata. Kitty clearly does not like this, so I'm going to put her down now. And the kitty lactates and gives birth to young like a cow, instead of laying eggs like a chicken, and they have fur and three special tiny bones in their ears that only mammals have, so they're in the class Mammalia. So, she is more closely related to a cow than a chicken. Good to know! And like a bunch of other placental mammals that eat meat like weasels (the mustelids), and dogs, (the canines), kitties are in the order Carnivora. And they're in the cat family, Felidae, whose members have lithe bodies and roundish heads and, except for cheetahs, retractable claws. And they're littler than tigers and panthers, which puts them in the genus Felis. And then, at the level of the species, the descriptions get pretty dang detailed, so let's just say that, you know what a cat is right? So the species name is catus. And look at that: Felis catus! Aw. Kitty. I could have that whole thing cross-stitched onto a pillow for you to sleep on! And it would be cute! Thank you for watching our taxider- I mean, our taxonomy episode of Crash Course Biology. We hope that you learned something. Thanks to everybody who helped put this episode together. If you have any questions for us, please leave them on Facebook or Twitter or in the comments below. And we will get to them. Hopefully very quickly. I will see you next time!
History
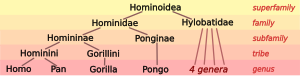
Human taxonomy on one hand involves the placement of humans within the taxonomy of the hominids (great apes), and on the other the division of archaic and modern humans into species and, if applicable, subspecies. Modern zoological taxonomy was developed by Carl Linnaeus during the 1730s to 1750s. He was the first to develop the idea that, like other biological entities, groups of people could too share taxonomic classifications.[7] He named the human species as Homo sapiens in 1758, as the only member species of the genus Homo, divided into several subspecies corresponding to the great races. The Latin noun homō (genitive hominis) means "human being". The systematic name Hominidae for the family of the great apes was introduced by John Edward Gray (1825).[8] Gray also supplied Hominini as the name of the tribe including both chimpanzees (genus Pan) and humans (genus Homo).
The discovery of the first extinct archaic human species from the fossil record dates to the mid 19th century: Homo neanderthalensis, classified in 1864. Since then, a number of other archaic species have been named, but there is no universal consensus as to their exact number. After the discovery of H. neanderthalensis, which even if "archaic" is recognizable as clearly human, late 19th to early 20th century anthropology for a time was occupied with finding the supposedly "missing link" between Homo and Pan. The "Piltdown Man" hoax of 1912 was the fraudulent presentation of such a transitional species. Since the mid-20th century, knowledge of the development of Hominini has become much more detailed, and taxonomical terminology has been altered a number of times to reflect this.
The introduction of Australopithecus as a third genus, alongside Homo and Pan, in the tribe Hominini is due to Raymond Dart (1925). Australopithecina as a subtribe containing Australopithecus as well as Paranthropus (Broom 1938) is a proposal by Gregory & Hellman (1939). More recently proposed additions to the Australopithecina subtribe include Ardipithecus (1995) and Kenyanthropus (2001). The position of Sahelanthropus (2002) relative to Australopithecina within Hominini is unclear. Cela-Conde and Ayala (2003) propose the recognition of Australopithecus, Ardipithecus, Praeanthropus, and Sahelanthropus (the latter incertae sedis)as separate genera.[9]
Other proposed genera, now mostly considered part of Homo, include: Pithecanthropus (Dubois, 1894), Protanthropus (Haeckel, 1895), Sinanthropus (Black, 1927), Cyphanthropus (Pycraft, 1928) Africanthropus (Dreyer, 1935),[10] Telanthropus (Broom & Anderson 1949), Atlanthropus (Arambourg, 1954), Tchadanthropus (Coppens, 1965).
The genus Homo has been taken to originate some two million years ago, since the discovery of stone tools in Olduvai Gorge, Tanzania, in the 1960s. Homo habilis (Leakey et al., 1964) would be the first "human" species (member of genus Homo) by definition, its type specimen being the OH 7 fossils. However, the discovery of more fossils of this type has opened up the debate on the delineation of H. habilis from Australopithecus. Especially, the LD 350-1 jawbone fossil discovered in 2013, dated to 2.8 Mya, has been argued as being transitional between the two.[11] It is also disputed whether H. habilis was the first hominin to use stone tools, as Australopithecus garhi, dated to c. 2.5 Mya, has been found along with stone tool implements.[12] Fossil KNM-ER 1470 (discovered in 1972, designated Pithecanthropus rudolfensis by Alekseyev 1978) is now seen as either a third early species of Homo (alongside H. habilis and H. erectus) at about 2 million years ago, or alternatively as transitional between Australopithecus and Homo.[13]
Wood and Richmond (2000) proposed that Gray's tribe Hominini ("hominins") be designated as comprising all species after the chimpanzee–human last common ancestor by definition, to the inclusion of Australopithecines and other possible pre-human or para-human species (such as Ardipithecus and Sahelanthropus) not known in Gray's time.[14] In this suggestion, the new subtribe of Hominina was to be designated as including the genus Homo exclusively, so that Hominini would have two subtribes, Australopithecina and Hominina, with the only known genus in Hominina being Homo. Orrorin (2001) has been proposed as a possible ancestor of Hominina but not Australopithecina.[15]
Designations alternative to Hominina have been proposed: Australopithecinae (Gregory & Hellman 1939) and Preanthropinae (Cela-Conde & Altaba 2002);[16]
Species
At least a dozen species of Homo other than Homo sapiens have been proposed, with varying degrees of consensus. Homo erectus is widely recognized as the species directly ancestral to Homo sapiens.[citation needed] Most other proposed species are proposed as alternatively belonging to either Homo erectus or Homo sapiens as a subspecies. This concerns Homo ergaster in particular.[17][18] One proposal divides Homo erectus into an African and an Asian variety; the African is Homo ergaster, and the Asian is Homo erectus sensu stricto. (Inclusion of Homo ergaster with Asian Homo erectus is Homo erectus sensu lato.)[19] There appears to be a recent trend, with the availability of ever more difficult-to-classify fossils such as the Dmanisi skulls (2013) or Homo naledi fossils (2015) to subsume all archaic varieties under Homo erectus.[20][21][22]
| Lineages | Temporal range (kya) |
Habitat | Adult height | Adult mass | Cranial capacity (cm3) |
Fossil record | Discovery | Publication of name |
|---|---|---|---|---|---|---|---|---|
| H. habilis membership in Homo uncertain |
2,100–1,500[a][b] | Tanzania | 110–140 cm (3 ft 7 in – 4 ft 7 in) | 33–55 kg (73–121 lb) | 510–660 | Many | 1960 | 1964 |
| H. rudolfensis membership in Homo uncertain |
1,900 | Kenya | 700 | 2 sites | 1972 | 1986 | ||
| H. gautengensis also classified as H. habilis |
1,900–600 | South Africa | 100 cm (3 ft 3 in) | 3 individuals[25][c] | 2010 | 2010 | ||
| H. erectus | 1,900–140[26][d][27][e] | Africa, Eurasia | 180 cm (5 ft 11 in) | 60 kg (130 lb) | 850 (early) – 1,100 (late) | Many[f][g] | 1891 | 1892 |
| H. ergaster African H. erectus |
1,800–1,300[29] | East and Southern Africa | 700–850 | Many | 1949 | 1975 | ||
| H. antecessor | 1,200–800 | Western Europe | 175 cm (5 ft 9 in) | 90 kg (200 lb) | 1,000 | 2 sites | 1994 | 1997 |
| H. heidelbergensis early H. neanderthalensis |
600–300[h] | Europe, Africa | 180 cm (5 ft 11 in) | 90 kg (200 lb) | 1,100–1,400 | Many | 1907 | 1908 |
| H. cepranensis a single fossil, possibly H. heidelbergensis |
c. 450[30] | Italy | 1,000 | 1 skull cap | 1994 | 2003 | ||
| H. longi | 309–138[31] | Northeast China | 1,420[32] | 1 individual | 1933 | 2021 | ||
| H. rhodesiensis early H. sapiens |
c. 300 | Zambia | 1,300 | Single or very few | 1921 | 1921 | ||
| H. naledi | c. 300[33] | South Africa | 150 cm (4 ft 11 in) | 45 kg (99 lb) | 450 | 15 individuals | 2013 | 2015 |
| H. sapiens (anatomically modern humans) |
c. 300–present[i] | Worldwide | 150–190 cm (4 ft 11 in – 6 ft 3 in) | 50–100 kg (110–220 lb) | 950–1,800 | (extant) | —— | 1758 |
| H. neanderthalensis |
240–40[36][j] | Europe, Western Asia | 170 cm (5 ft 7 in) | 55–70 kg (121–154 lb) (heavily built) |
1,200–1,900 | Many | 1829 | 1864 |
| H. floresiensis classification uncertain |
190–50 | Indonesia | 100 cm (3 ft 3 in) | 25 kg (55 lb) | 400 | 7 individuals | 2003 | 2004 |
| Nesher Ramla Homo classification uncertain |
140–120 | Israel | several individuals | 2021 | ||||
| H. tsaichangensis possibly H. erectus or Denisova |
c. 100[k] | Taiwan | 1 individual | 2008(?) | 2015 | |||
| H. luzonensis |
c. 67[39][40] | Philippines | 3 individuals | 2007 | 2019 | |||
| Denisova hominin | 40 | Siberia | 2 sites | 2000 |
2010[l] |
Subspecies
Homo sapiens subspecies

The recognition or nonrecognition of subspecies of Homo sapiens has a complicated history. The rank of subspecies in zoology is introduced for convenience, and not by objective criteria, based on pragmatic consideration of factors such as geographic isolation and sexual selection. The informal taxonomic rank of race is variously considered equivalent or subordinate to the rank of subspecies, and the division of anatomically modern humans (H. sapiens) into subspecies is closely tied to the recognition of major racial groupings based on human genetic variation.
A subspecies cannot be recognized independently: a species will either be recognized as having no subspecies at all or at least two (including any that are extinct). Therefore, the designation of an extant subspecies Homo sapiens sapiens only makes sense if at least one other subspecies is recognized. H. s. sapiens is attributed to "Linnaeus (1758)" by the taxonomic Principle of Coordination.[42] During the 19th to mid-20th century, it was common practice to classify the major divisions of extant H. sapiens as subspecies, following Linnaeus (1758), who had recognized H. s. americanus, H. s. europaeus, H. s. asiaticus and H. s. afer as grouping the native populations of the Americas, West Eurasia, East Asia and Sub-Saharan Africa, respectively. Linnaeus also included H. s. ferus, for the "wild" form which he identified with feral children, and two other "wild" forms for reported specimens now considered very dubious (see cryptozoology), H. s. monstrosus and H. s. troglodytes.[43]
There were variations and additions to the categories of Linnaeus, such as H. s. tasmanianus for the native population of Australia.[44] Bory de St. Vincent in his Essai sur l'Homme (1825) extended Linnaeus's "racial" categories to as many as fifteen: Leiotrichi ("smooth-haired"): japeticus (with subraces), arabicus, iranicus, indicus, sinicus, hyperboreus, neptunianus, australasicus, columbicus, americanus, patagonicus; Oulotrichi ("crisp-haired"): aethiopicus, cafer, hottentotus, melaninus.[45] Similarly, Georges Vacher de Lapouge (1899) also had categories based on race, such as priscus, spelaeus (etc.).
Homo sapiens neanderthalensis was proposed by King (1864) as an alternative to Homo neanderthalensis.[46] There have been "taxonomic wars" over whether Neanderthals were a separate species since their discovery in the 1860s. Pääbo (2014) frames this as a debate that is unresolvable in principle, "since there is no definition of species perfectly describing the case."[47] Louis Lartet (1869) proposed Homo sapiens fossilis based on the Cro-Magnon fossils.
There are a number of proposals of extinct varieties of Homo sapiens made in the 20th century. Many of the original proposals were not using explicit trinomial nomenclature, even though they are still cited as valid synonyms of H. sapiens by Wilson & Reeder (2005).[48] These include: Homo grimaldii (Lapouge, 1906), Homo aurignacensis hauseri (Klaatsch & Hauser, 1910), Notanthropus eurafricanus (Sergi, 1911), Homo fossilis infrasp. proto-aethiopicus (Giuffrida-Ruggeri, 1915), Telanthropus capensis (Broom, 1917),[49] Homo wadjakensis (Dubois, 1921), Homo sapiens cro-magnonensis, Homo sapiens grimaldiensis (Gregory, 1921), Homo drennani (Kleinschmidt, 1931),[50] Homo galilensis (Joleaud, 1931) = Paleanthropus palestinus (McCown & Keith, 1932).[51] Rightmire (1983) proposed Homo sapiens rhodesiensis.[52]
After World War II, the practice of dividing extant populations of Homo sapiens into subspecies declined. An early authority explicitly avoiding the division of H. sapiens into subspecies was Grzimeks Tierleben, published 1967–1972.[53] A late example of an academic authority proposing that the human racial groups should be considered taxonomical subspecies is John Baker (1974).[54] The trinomial nomenclature Homo sapiens sapiens became popular for "modern humans" in the context of Neanderthals being considered a subspecies of H. sapiens in the second half of the 20th century. Derived from the convention, widespread in the 1980s, of considering two subspecies, H. s. neanderthalensis and H. s. sapiens, the explicit claim that "H. s. sapiens is the only extant human subspecies" appears in the early 1990s.[55]
Since the 2000s, the extinct Homo sapiens idaltu (White et al., 2003) has gained wide recognition as a subspecies of Homo sapiens, but even in this case there is a dissenting view arguing that "the skulls may not be distinctive enough to warrant a new subspecies name".[56] H. s. neanderthalensis and H. s. rhodesiensis continue to be considered separate species by some authorities, but the 2010s discovery of genetic evidence of archaic human admixture with modern humans has reopened the details of taxonomy of archaic humans.[57]
Homo erectus subspecies
Homo erectus since its introduction in 1892 has been divided into numerous subspecies, many of them formerly considered individual species of Homo. None of these subspecies have universal consensus among paleontologists.
- Homo erectus erectus (Java Man) (1970s)[58]
- Homo erectus yuanmouensis (Yuanmou Man) (Li et al., 1977)
- Homo erectus lantianensis (Lantian Man) (Woo Ju-Kang, 1964)
- Homo erectus nankinensis (Nanjing Man) (1993)
- Homo erectus pekinensis (Peking Man) (1970s)[58]
- Homo erectus palaeojavanicus (Meganthropus) (Tyler, 2001)
- Homo erectus soloensis (Solo Man) (Oppenoorth, 1932)
- Homo erectus tautavelensis (Tautavel Man) (de Lumley and de Lumley, 1971)
- Homo erectus georgicus (1991)
- Homo erectus bilzingslebenensis (Vlček, 2002)[59]
See also
Footnotes
- ^ Confirmed H. habilis fossils are dated to between 2.1 and 1.5 million years ago. This date range overlaps with the emergence of Homo erectus.[23][24]
- ^ Hominins with "proto-Homo" traits may have lived as early as 2.8 million years ago, as suggested by a fossil jawbone classified as transitional between Australopithecus and Homo discovered in 2015.
- ^ A species proposed in 2010 based on the fossil remains of three individuals dated between 1.9 and 0.6 million years ago. The same fossils were also classified as H. habilis, H. ergaster or Australopithecus by other anthropologists.
- ^ H. erectus may have appeared some 2 million years ago. Fossils dated to as much as 1.8 million years ago have been found both in Africa and in Southeast Asia, and the oldest fossils by a narrow margin (1.85 to 1.77 million years ago) were found in the Caucasus, so that it is unclear whether H. erectus emerged in Africa and migrated to Eurasia, or if, conversely, it evolved in Eurasia and migrated back to Africa.
- ^ Homo erectus soloensis, found in Java, is considered the latest known survival of H. erectus. Formerly dated to as late as 50,000 to 40,000 years ago, a 2011 study pushed back the date of its extinction of H. e. soloensis to 143,000 years ago at the latest, more likely before 550,000 years ago. [28]
- ^ Now also included in H. erectus are Peking Man (formerly Sinanthropus pekinensis) and Java Man (formerly Pithecanthropus erectus).
- ^ H. erectus is now grouped into various subspecies, including Homo erectus erectus, Homo erectus yuanmouensis, Homo erectus lantianensis, Homo erectus nankinensis, Homo erectus pekinensis, Homo erectus palaeojavanicus, Homo erectus soloensis, Homo erectus tautavelensis, Homo erectus georgicus. The distinction from descendant species such as Homo ergaster, Homo floresiensis, Homo antecessor, Homo heidelbergensis and indeed Homo sapiens is not entirely clear.
- ^ The type fossil is Mauer 1, dated to ca. 0.6 million years ago. The transition from H. heidelbergensis to H. neanderthalensis between 300 and 243 thousand years ago is conventional, and makes use of the fact that there is no known fossil in this period. Examples of H. heidelbergensis are fossils found at Bilzingsleben (also classified as Homo erectus bilzingslebensis).
- ^ The age of H. sapiens has long been assumed to be close to 200,000 years, but since 2017 there have been a number of suggestions extending this time to as high as 300,000 years. In 2017, fossils found in Jebel Irhoud (Morocco) suggest that Homo sapiens may have speciated by as early as 315,000 years ago.[34] Genetic evidence has been adduced for an age of roughly 270,000 years.[35]
- ^ The first humans with "proto-Neanderthal traits" lived in Eurasia as early as 0.6 to 0.35 million years ago (classified as H. heidelbergensis, also called a chronospecies because it represents a chronological grouping rather than being based on clear morphological distinctions from either H. erectus or H. neanderthalensis). There is a fossil gap in Europe between 300 and 243 kya, and by convention, fossils younger than 243 kya are called "Neanderthal".[37]
- ^ younger than 450 kya, either between 190–130 or between 70–10 kya[38]
- ^ provisional names Homo sp. Altai or Homo sapiens ssp. Denisova.
References
- ^ Stringer, Chris (June 12, 2003). "Human evolution: Out of Ethiopia". Nature. 423 (6941): 693–695. Bibcode:2003Natur.423..692S. doi:10.1038/423692a. PMID 12802315. S2CID 26693109.
- ^ "Herto skulls (Homo sapiens idaltu)". talkorigins org. Retrieved June 7, 2016.
- ^ Stringer, C. (2016). "The origin and evolution of Homo sapiens". Philosophical Transactions of the Royal Society B: Biological Sciences. 371 (1698): 20150237. doi:10.1098/rstb.2015.0237. PMC 4920294. PMID 27298468.
- ^ Jared Diamond in The Third Chimpanzee (1991), and Morris Goodman (2003) Hecht, Jeff (May 19, 2003). "Chimps are human, gene study implies". New Scientist. Retrieved December 8, 2011.
- ^ K. Wagner, Jennifer (2016). "Anthropologists' views on race, ancestry, and genetics". American Journal of Physical Anthropology. 162 (2): 318–327. doi:10.1002/ajpa.23120. PMC 5299519. PMID 27874171.
- ^ "AAA Statement on Race". American Anthropological Association.
- ^ Marks, Jonathan (May 2007). "Long shadow of Linnaeus's human taxonomy". Nature. 447 (7140): 28. Bibcode:2007Natur.447Q..28M. doi:10.1038/447028a. ISSN 1476-4687. PMID 17476243. S2CID 33066198.
- ^ J. E. Gray, "An outline of an attempt at the disposition of Mammalia into Tribes and Families, with a list of genera apparently appertaining to each Tribe", Annals of Philosophy, new series (1825), pp. 337–344.
- ^ Cela-Conde, C. J.; Ayala, F. J. (2003). "Genera of the human lineage". Proceedings of the National Academy of Sciences. 100 (13): 7684–7689. Bibcode:2003PNAS..100.7684C. doi:10.1073/pnas.0832372100. PMC 164648. PMID 12794185.
- ^ Introduced for the Florisbad Skull (discovered in 1932, Homo florisbadensis or Homo helmei). Also the genus suggested for a number of archaic human skulls found at Lake Eyasi by Weinert (1938). Leaky, Journal of the East Africa Natural History Society (1942), p. 43.
- ^ Villmoare, B. (2015). "Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia". Science. 347 (6228): 1352–1355. Bibcode:2015Sci...347.1352V. doi:10.1126/science.aaa1343. PMID 25739410.. Some paleoanthropologists regard the H. habilis taxon as invalid, made up of fossil specimens of Australopithecus and Homo. Tattersall, I. & Schwartz, J.H., Extinct Humans, Westview Press, New York, 2001, p. 111.
- ^ De Heinzelin, J; Clark, JD; White, T; Hart, W; Renne, P; Woldegabriel, G; Beyene, Y; Vrba, E (1999). "Environment and behavior of 2.5-million-year-old Bouri hominids". Science. 284 (5414): 625–9. Bibcode:1999Sci...284..625D. doi:10.1126/science.284.5414.625. PMID 10213682.
- ^ Kaplan, Matt (August 8, 2012). "Fossils point to a big family for human ancestors". Nature. doi:10.1038/nature.2012.11144. S2CID 87482930. Retrieved August 8, 2012.
- ^ Wood, Bernard; Richmond, Brian G. (2000). "Human evolution: taxonomy and paleobiology". Journal of Anatomy. 197 (Pt 1): 19–60. doi:10.1046/j.1469-7580.2000.19710019.x. PMC 1468107. PMID 10999270.
- ^ Reynolds, Sally C; Gallagher, Andrew (March 29, 2012). African Genesis: Perspectives on Hominin Evolution. Cambridge University Press. ISBN 9781107019959.
- ^ Brunet, M.; et al. (2002). "A new hominid from the upper Miocene of Chad, central Africa" (PDF). Nature. 418 (6894): 145–151. Bibcode:2002Natur.418..145B. doi:10.1038/nature00879. PMID 12110880. S2CID 1316969. Cela-Conde, C.J.; Ayala, F.J. (2003). "Genera of the human lineage". PNAS. 100 (13): 7684–7689. Bibcode:2003PNAS..100.7684C. doi:10.1073/pnas.0832372100. PMC 164648. PMID 12794185. Wood, B.; Lonergan, N. (2008). "The hominin fossil record: taxa, grades and clades" (PDF). Journal of Anatomy. 212 (4): 354–376. doi:10.1111/j.1469-7580.2008.00871.x. PMC 2409102. PMID 18380861.
- ^ Hazarika, Manji (16–30 June 2007). "Homo erectus / ergaster and Out of Africa: Recent Developments in Paleoanthropology and Prehistoric Archaeology" (PDF).
- ^ Klein, R. (1999). The Human Career: Human biological and cultural origins. Chicago, IL: University of Chicago Press. ISBN 0226439631.
- ^ Antón, S.C. (2003). "Natural history of Homo erectus". American Journal of Physical Anthropology. 122: 126–170. doi:10.1002/ajpa.10399. PMID 14666536.
By the 1980s, the growing numbers of H. erectus specimens, particularly in Africa, led to the realization that Asian H. erectus (H. erectus sensu stricto), once thought so primitive, was in fact more derived than its African counterparts. These morphological differences were interpreted by some as evidence that more than one species might be included in H. erectus sensu lato (e.g., Stringer, 1984; Andrews, 1984; Tattersall, 1986; Wood, 1984, 1991a, b; Schwartz and Tattersall, 2000) ... Unlike the European lineage, in my opinion, the taxonomic issues surrounding Asian vs. African H. erectus are more intractable. The issue was most pointedly addressed with the naming of H. ergaster on the basis of the type mandible KNM-ER 992, but also including the partial skeleton and isolated teeth of KNM-ER 803 among other Koobi Fora remains (Groves and Mazak, 1975). Recently, this specific name was applied to most early African and Georgian H. erectus in recognition of the less-derived nature of these remains vis à vis conditions in Asian H. erectus (see Wood, 1991a, p. 268; Gabunia et al., 2000a). At least portions of the paratype of H. ergaster (e.g., KNM-ER 1805) are not included in most current conceptions of that taxon. The H. ergaster question remains famously unresolved (e.g., Stringer, 1984; Tattersall, 1986; Wood, 1991a, 1994; Rightmire, 1998b; Gabunia et al., 2000a; Schwartz and Tattersall, 2000), in no small part because the original diagnosis provided no comparison with the Asian fossil record.
- ^ "Skull suggests three early human species were one". News & Comment. Nature.
- ^ Lordkipanidze, David; Ponce de Leòn, Marcia S.; Margvelashvili, Ann; Rak, Yoel; Rightmire, G. Philip; Vekua, Abesalom; Zollikofer, Christoph P. E. (18 October 2013). "A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo". Science. 342 (6156): 326–331. Bibcode:2013Sci...342..326L. doi:10.1126/science.1238484. PMID 24136960. S2CID 20435482.
- ^ Switek, Brian (17 October 2013). "Beautiful skull spurs debate on human history". National Geographic. Archived from the original on October 17, 2013. Retrieved 22 September 2014.
- ^ Schrenk F, Kullmer O, Bromage T (2007). "The Earliest Putative Homo Fossils". In Henke W, Tattersall I (eds.). Handbook of Paleoanthropology. Vol. 1. In collaboration with Thorolf Hardt. Berlin, Heidelberg: Springer. pp. 1611–1631. doi:10.1007/978-3-540-33761-4_52. ISBN 978-3-540-32474-4.
- ^ DiMaggio EN, Campisano CJ, Rowan J, Dupont-Nivet G, Deino AL, Bibi F, et al. (March 2015). "Paleoanthropology. Late Pliocene fossiliferous sedimentary record and the environmental context of early Homo from Afar, Ethiopia". Science. 347 (6228): 1355–9. Bibcode:2015Sci...347.1355D. doi:10.1126/science.aaa1415. PMID 25739409. S2CID 43455561.
- ^ Curnoe D (June 2010). "A review of early Homo in southern Africa focusing on cranial, mandibular and dental remains, with the description of a new species (Homo gautengensis sp. nov.)". Homo. 61 (3): 151–77. doi:10.1016/j.jchb.2010.04.002. PMID 20466364.
- ^ Haviland WA, Walrath D, Prins HE, McBride B (2007). Evolution and Prehistory: The Human Challenge (8th ed.). Belmont, CA: Thomson Wadsworth. p. 162. ISBN 978-0-495-38190-7.
- ^ Ferring R, Oms O, Agustí J, Berna F, Nioradze M, Shelia T, et al. (June 2011). "Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85-1.78 Ma". Proceedings of the National Academy of Sciences of the United States of America. 108 (26): 10432–6. Bibcode:2011PNAS..10810432F. doi:10.1073/pnas.1106638108. PMC 3127884. PMID 21646521.
- ^ Indriati E, Swisher CC, Lepre C, Quinn RL, Suriyanto RA, Hascaryo AT, et al. (2011). "The age of the 20 meter Solo River terrace, Java, Indonesia and the survival of Homo erectus in Asia". PLOS ONE. 6 (6): e21562. Bibcode:2011PLoSO...621562I. doi:10.1371/journal.pone.0021562. PMC 3126814. PMID 21738710.
- ^ Hazarika M (2007). "Homo erectus/ergaster and Out of Africa: Recent Developments in Paleoanthropology and Prehistoric Archaeology" (PDF). EAA Summer School eBook. Vol. 1. European Anthropological Association. pp. 35–41.
Intensive Course in Biological Anthrpology, 1st Summer School of the European Anthropological Association, 16–30 June, 2007, Prague, Czech Republic
- ^ Muttoni G, Scardia G, Kent DV, Swisher CC, Manzi G (2009). "Pleistocene magnetochronology of early hominin sites at Ceprano and Fontana Ranuccio, Italy". Earth and Planetary Science Letters. 286 (1–2): 255–268. Bibcode:2009E&PSL.286..255M. doi:10.1016/j.epsl.2009.06.032.
- ^ Ji Q, Wu W, Ji Y, Li Q, Ni X (25 June 2021). "Late Middle Pleistocene Harbin cranium represents a new Homo species". The Innovation. 2 (3): 100132. doi:10.1016/j.xinn.2021.100132. PMC 8454552. PMID 34557772.
- ^ Ni X, Ji Q, Wu W, Shao Q, Ji Y, Zhang C, Liang L, Ge J, Guo Z, Li J, Li Q, Grün R, Stringer C (25 June 2021). "Massive cranium from Harbin in northeastern China establishes a new Middle Pleistocene human lineage". The Innovation. 2 (3): 100130. doi:10.1016/j.xinn.2021.100130. PMC 8454562. PMID 34557770.
- ^ Dirks PH, Roberts EM, Hilbert-Wolf H, Kramers JD, Hawks J, Dosseto A, et al. (May 2017). "Homo naledi and associated sediments in the Rising Star Cave, South Africa". eLife. 6: e24231. doi:10.7554/eLife.24231. PMC 5423772. PMID 28483040.
- ^ Callaway, Ewan (7 June 2017). "Oldest Homo sapiens fossil claim rewrites our species' history". Nature. doi:10.1038/nature.2017.22114. Retrieved 11 June 2017.
- ^ Posth C, Wißing C, Kitagawa K, Pagani L, van Holstein L, Racimo F, et al. (July 2017). "Deeply divergent archaic mitochondrial genome provides lower time boundary for African gene flow into Neanderthals". Nature Communications. 8: 16046. Bibcode:2017NatCo...816046P. doi:10.1038/ncomms16046. PMC 5500885. PMID 28675384.
- ^ Bischoff JL, Shamp DD, Aramburu A, et al. (March 2003). "The Sima de los Huesos Hominids Date to Beyond U/Th Equilibrium (>350 kyr) and Perhaps to 400–500 kyr: New Radiometric Dates". Journal of Archaeological Science. 30 (3): 275–280. doi:10.1006/jasc.2002.0834. ISSN 0305-4403.
- ^ Dean D, Hublin JJ, Holloway R, Ziegler R (May 1998). "On the phylogenetic position of the pre-Neandertal specimen from Reilingen, Germany". Journal of Human Evolution. 34 (5): 485–508. doi:10.1006/jhev.1998.0214. PMID 9614635.
- ^ Chang CH, Kaifu Y, Takai M, Kono RT, Grün R, Matsu'ura S, et al. (January 2015). "The first archaic Homo from Taiwan". Nature Communications. 6: 6037. Bibcode:2015NatCo...6.6037C. doi:10.1038/ncomms7037. PMC 4316746. PMID 25625212.
- ^ Détroit F, Mijares AS, Corny J, Daver G, Zanolli C, Dizon E, et al. (April 2019). "A new species of Homo from the Late Pleistocene of the Philippines" (PDF). Nature. 568 (7751): 181–186. Bibcode:2019Natur.568..181D. doi:10.1038/s41586-019-1067-9. PMID 30971845. S2CID 106411053.
- ^ Zimmer C (10 April 2019). "A new human species once lived in this Philippine cave – Archaeologists in Luzon Island have turned up the bones of a distantly related species, Homo luzonensis, further expanding the human family tree". The New York Times. Retrieved 10 April 2019.
- ^ Ralph, Bob (February 19, 1987). "Conforming to type". New Scientist. No. 1548. p. 59.
as far as I know, there is no type material for Homo sapiens. To be fair to Linnaeus, the practice of setting type specimens aside doesn't seem to have developed until a century or so later.
- ^ "article 46.1". ICZN glossary (4th ed.). International Code of Zoological Nomenclature. Archived from the original on 2016-03-03. Retrieved 2018-06-04.
Statement of the Principle of Coordination applied to species-group names. A name established for a taxon at either rank in the species group is deemed to have been simultaneously established by the same author for a taxon at the other rank in the group; both nominal taxa have the same name-bearing type, whether that type was fixed originally or subsequently.
Homo sapiens sapiens is rarely used before the 1940s. In 1946, John Wendell Bailey attributes the name to Linnaeus (1758) explicitly: "Linnaeus. Syst. Nat. ed. 10, Vol. 1. pp. 20, 21, 22, lists five races of man, viz: Homo sapiens sapiens (white — Caucasian) [...]", This is a misattribution, but H. s. sapiens has since often been attributed to Linnaeus. In actual fact, Linnaeus, Syst. Nat. ed. 10 Vol. 1. p. 21 does not have Homo sapiens sapiens, the "white" or "Caucasian" race being instead called Homo sapiens Europaeus. This is explicitly pointed out in Bulletin der Schweizerische Gesellschaft für Anthropologie und Ethnologie Volume 21 (1944), p. 18 (arguing not against H. s. sapiens but against "H. s. albus L." proposed by von Eickstedt and Peters): "die europide Rassengruppe, als Subspecies aufgefasst, [würde] Homo sapiens eurpoaeus L. heissen" ("the Europid racial group, considered as a subspecies, would be named H. s. europeaeus L."). See also: John R. Baker, Race, Oxford University Press (1974), 205. - ^ Linné, Carl von (1758). Systema naturæ. Regnum animale (10 ed.). pp. 18ff.
- ^ See e.g. John Wendell Bailey, The Mammals of Virginia (1946), p. 356.; Journal of Mammalogy 26-27 (1945), p. 359.; J. Desmond Clark (ed.), The Cambridge History of Africa, Cambridge University Press (1982), p. 141 (with references).
- ^ Annals of Philosophy 11, London (1826), p. 71
- ^ Frederick S. Szalay, Eric Delson, Evolutionary History of the Primates (2013), 508
- ^ Pääbo, Svante (2014). Neanderthal Man: In Search of Lost Genomes. New York: Basic Books. p. 237.
- ^ Groves, C. P. (2005). Wilson, D. E.; Reeder, D. M. (eds.). Mammal Species of the World: A Taxonomic and Geographic Reference (3rd ed.). Baltimore: Johns Hopkins University Press. ISBN 0-801-88221-4. OCLC 62265494.
- ^ Harrison, T. (2013). Kimbel, William H.; Martin, Lawrence B. (eds.). Species, Species Concepts and Primate Evolution. Springer. p. 361. ISBN 9781489937452.
- ^ M. R. Drennan, "An Australoid Skull from the Cape Flats", The Journal of the Royal Anthropological Institute of Great Britain and Ireland Vol. 59 (Jul. - Dec., 1929), 417-427.
- ^ among other names suggested for fossils later subsumed under neanderthalensis, see: Delson, Eric; Tattersall, Ian; Couvering, John Van; Brooks, Alison S. (2004). Encyclopedia of Human Evolution and Prehistory (Second ed.). Routledge. ISBN 9781135582272.
- ^ Rightmire, GP (June 3, 1983). "The Lake Ndutu cranium and early Homo sapiens in Africa". American Journal of Physical Anthropology. 61 (2): 245–54. doi:10.1002/ajpa.1330610214. PMID 6410925.
- ^ English translation (1972–1975): Grzimek's Animal Life Encyclopedia, Volume 11, p. 55.
- ^ John R. Baker, Race, Oxford University Press (1974).
- ^ Kitahara, Michio (1991). The tragedy of evolution: the human animal confronts modern society. p. xi.
We are the only surviving subspecies of Homo sapiens
- ^ Stringer, Chris (June 12, 2003). "Human evolution: Out of Ethiopia". Nature. 423 (6941): 692–3, 695. Bibcode:2003Natur.423..692S. doi:10.1038/423692a. PMID 12802315. S2CID 26693109.
- ^ Hublin, J. J. (2009). "The origin of Neandertals". Proceedings of the National Academy of Sciences. 106 (38): 16022–7. Bibcode:2009PNAS..10616022H. doi:10.1073/pnas.0904119106. JSTOR 40485013. PMC 2752594. PMID 19805257. Harvati, K.; Frost, S.R.; McNulty, K.P. (2004). "Neanderthal taxonomy reconsidered: implications of 3D primate models of intra- and interspecific differences". Proceedings of the National Academy of Sciences. 101 (5): 1147–52. Bibcode:2004PNAS..101.1147H. doi:10.1073/pnas.0308085100. PMC 337021. PMID 14745010. "Homo neanderthalensis King, 1864". Wiley-Blackwell Encyclopedia of Human Evolution. Chichester, West Sussex: Wiley-Blackwell. 2013. pp. 328–331.
- ^ a b In the 1970s a tendency developed to regard the Javanese variety of H. erectus as a subspecies, Homo erectus erectus, with the Chinese variety being referred to as Homo erectus pekinensis. See: Sartono, S. (12 May 2011). "Implications arising from Pithecanthropus VIII". In Tuttle, Russell H. (ed.). Paleoanthropology: Morphology and Paleoecology. Walter de Gruyter. p. 328. ISBN 9783110810691.
- ^ Emanuel Vlček: Der fossile Mensch von Bilzingsleben (= Bilzingsleben. Bd. 6 = Beiträge zur Ur- und Frühgeschichte Mitteleuropas 35). Beier & Beran, Langenweißbach 2002.
| Taxonomy (Hominins) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ancestors |
| ||||||||||||||||||||||||||
| Models |
| ||||||||||||||||||||||||||
| Timelines | |||||||||||||||||||||||||||
| Others | |||||||||||||||||||||||||||